

1. 서론
2. 재료 및 방법
2.1 환경요인
2.2 중형동물플랑크톤 채집과 동정
2.3 자료분석
3. 결과
3.1 환경요인
3.2 동물플랑크톤의 종조성
3.3 요각류의 출현양상과 주요 우점종
3.4 우점 출현하는 요각류와 환경과의 상관 관계 및 집괴분석
4. 고찰
1. 서론
강 하구는 육상에서 유입되는 담수와 해수가 만나는 해역으로 물리 및 화학적 조건은 시공간적으로 매우 다양하며, 염분 구배는 하구 생태계 구조 변화에 중요 요인으로 작용한다(Martino and Able 2003; Selleslagh and Amara 2008). 하구생태계는 육상에서 공급되는 에너지원의 유입으로 결정되며, 이 에너지는 물의 흐름과 수문, 하구둑에 영향을 받는다(Jekins and Boulton 2003). 그러나 방조제의 건설은 하구로 유입되는 강물의 흐름을 방해하여 하구 생태계에 커다란 변화를 초래한다. 특히, 하구에 서식하는 동물플랑크톤은 염분 구배에 따라 출현 특성을 달리하기 때문에, 하구둑 수문 조절에 의한 인위적인 담수의 유입은 하구 동물플랑크톤 군집 구조를 크게 변화시킬 수 있다(Mallin 1991). 이외도 하구둑은 산란장으로 회유하는 어류의 이동을 막거나 지연시켜 종의 쇠퇴 및 멸종에 영향을 미친다(Alhassan et al. 2015).
특히 네델란드 Oosterschelde 하구의 경우 해일 방지벽(storm-surge barrier) 건설 진행 중에도 하구 내 동물플랑크톤의 종조성과 생체량 변화가 발생하였다(Bakker 1994; Bakker and Rijswijk 1994). 우리나라의 경우에도 하구 매립뿐만 아니라 하구 방조제 건설이 하구 생태계의 플랑크톤 군집 변화에 크게 영향을 미칠 수 있음을 보고하고 있다(Lee et al. 2007; Lee et al. 2009). 특히, 새만금 방조제가 건설되기 전 만경강 하구에서는 기수종과 연안종이 출현하였으나, 방조제 건설 후 방조제 안에서는 기수성 요각류가 크게 증가한 반면, 바다 쪽에서는 연안에 출현하는 동물플랑크톤이 우점하였다(Suh et al. 1991; Lee et al. 2009).
한편 본 연구지역인 영산강 하구는 댐 건설로 인해 수문 개방에 의해 조절되는 일시적인 하구 기능을 회복하는 일시 개방형 하구로 변하였다. 일시 개방형 하구는 인위적인 수문조작에 의해 담수가 대량 유입될 때, 염분 변화가 일시에 큰 폭으로 변화할 뿐만 아니라 다량의 부유물질이 공급됨으로써 생태계의 교란을 초래하게 된다(Fisher et al. 1988). 실질적으로 인위적인 수문 조작으로 인한 담수 유입이 하구 해역에 영향을 미치는 연구는 일부 낙동강 하구역과 남대천 하구 등에서 조사된 바 있으나 그 사례가 많지 않다(Kang and Ihm 1998; Kwon et al. 2005). 따라서 본 연구는 영산강 방조제의 인위적인 수문 개폐에 의한 담수 유입이 해양 먹이사슬에서 일차생산자와 어패류 등의 상위소비자와의 중요한 연결고리 역할을 하고 있는 동물플랑크톤 군집 구조 변화에 미치는 영향을 살펴보고자 한다.
2. 재료 및 방법
2.1 환경요인
영산강 방조제의 수문 개방에 따른 담수 유입이 하구 내 환경인자의 변화와 동물플랑크톤 군집에 미치는 영향을 알아보기 위하여 2004년 2월부터 2006년 1월까지 영산강 하구역 8개 정점(하구 내 4개 정점, 하구 바깥 4개 정점)에서 조사를 실시하였다(Fig. 1). 이 중 2004년 6, 8, 9월, 2005년 4, 7월의 조사는 수문개방 후에 이루어졌으며, 이외의 조사는 수문이 폐쇄된 시기에 실시하였다. 따라서 이후의 서술부터는 2004년 6, 8, 9월, 2005년 4, 7월의 조사는 수문 개방 시기로, 이외의 조사는 수문 비개방 시기로 구분하여 서술하고자 한다.
강수량은 기상청(http://www.kma.go.kr)의 자료를 이용하였으며, 영산강 방조제의 담수 방류량은 한국농촌공사(http://www.ekr.or.kr)에서 실측된 일일 자료를 사용하여 월 간격으로 합계하였다.
조사해역의 해황 및 담수의 확산 정도를 파악하기 위해 수온과 염분은 YSI (Model 85 SCT)측정 장비를 이용하여 표층에서 1 m 아래의 수심과 저층에서 1 m 위를 측정하였다. 또한 엽록소 a 농도는 해수 100 ml를 유리 섬유 여과지로 필터 하였다. 여과된 필터들은 엽록소 a 추출 용액인 아세톤 90% 8 ml가 담긴 차광 시험관에 넣고 12시간이 지난 후, 24시간 내로 Fluorometer (Turner Designs 10-AU)를 사용하여 총 엽록소 a와 미소 엽록소 a농도를 측정하였다.
2.2 중형동물플랑크톤 채집과 동정
동물플랑크톤 채집은 NORPAC 네트(망목 220 μm, 망구직경 45 cm)를 이용하여 1 m/s의 속도로 3회 수직 채집하였다. 또한 정량분석을 위하여 망구에 유속계(General Oceanics Co., USA)를 부착하여 여과 해수량을 측정하였다. 채집된 시료는 선상에서 중성포르말린을 사용하여 시료의 최종농도가 5%가 되도록 고정하였다. 동물플랑크톤 종조성 및 출현 개체수 파악을 위해 시료의 분할이 필요한 경우 Folsom식 분할기를 이용하여 1/2~1/8까지 분할 후, UNESCO식 계수판에 옮긴 후 계수하였다. 동물플랑크톤 동정은 해부현미경(Nikon SMX 1000, Japan)을 사용하여 요각류를 중심으로 종 수준까지 동정하였다. 종 동정을 위해 보다 세밀한 관찰이 필요한 경우 종 동정을 특정할 수 있는 부속지를 해부하여 표본을 만든 후, 고배율 광학현미경(Nikon ECLIPSE 80i, Japan)을 이용하여 관찰하였다. 동물플랑크톤의 출현 개체수는 1 m3당 개체수로 환산하였다.
2.3 자료분석
수문 개폐에 따른 담수의 유입이 염분의 변동을 주어 동물플랑크톤의 종 다양성에 미치는 영향 정도를 평가하기 위하여 종다양성 지수(H')를 Shannon-Wiever (1963)의 다음 식에 의하여 구하였다.
s = 출현 종 수,
Pi = 총 출현 개체수에 대한 i종의 개체수 비율
또한 영산강 하구역을 조사시기, 지리적 위치, 수문의 개폐에 관계없이 Seine 하구역 구분에 따라 염분 범위만을 고려하여 저염분 지역(5 psu 이하), 중염분 지역(5~18 psu), 고염분 지역(18 psu 이상)으로 3개의 지역으로 구분하였다(Mouny and Dauvin 2002). 수문의 개폐에 의해 조사된 요각류의 출현 양상을 파악하기 위해서 요각류 개체수를 바탕으로 다변량 분석법인 분류법(classification)과 배열법(ordination)을 이용하여 군집분석을 수행하였다. 군집분석에 대한 분류군의 출현 개체수 자료는 조사시기와 종간의 심한 개체군 밀도차이에 의한 자료의 편중을 피하고 대상 자료의 분포를 정규화하기 위하여 log(x+1)로 변환한 자료를 이용하였다. 종 수준으로 동정된 요각류 군집의 정점 간 유사도 측정을 위해 Bray-Curtis 유사도 지수를 토대로 비가중 산술평균(UPGMA)에 의하여 군집화하는 계보적 집괴분석(hierarchical cluster analysis)을 시행하여 nMDS (non-metric multidimensional scaling) 배열법으로 군집분석을 수행하였다. 환경요인과 우점 요각류의 상호관계를 분석하기 위해 정준대응분석(Canonical Correspondence Anlaysis: CCA, CANOCO Ver4.5)을 사용하였다. 정준대응분석은 종조성과 환경변수들과의 관계를 이해하는데 널리 사용하는 다변량 통계분석법이다(ter Braak 1986).
3. 결과
3.1 환경요인
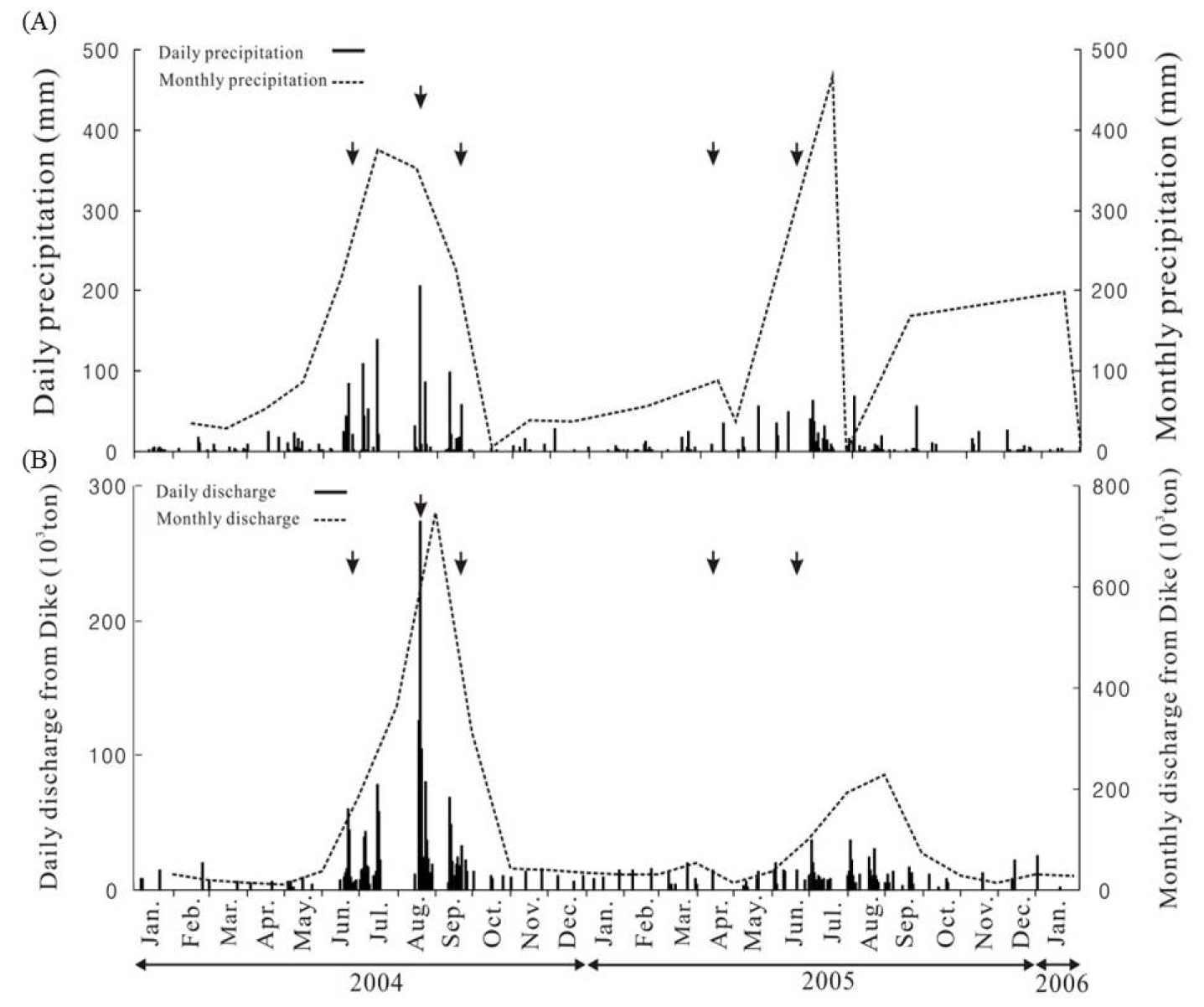
연구기간 동안 강우량 변화를 살펴보면, 장마 또는 태풍의 영향을 받는 하계에 평균 747 mm로 가장 높게 나타났으며, 동계에 평균 54.9 mm이하로 상대적으로 적은 강우량을 보였다(Fig. 2(A)). 강우량이 많은 하계에는 수문의 개방으로 월 평균 22,409 ton의 담수가 방류된 반면, 상대적으로 갈수기인 봄에는 월 평균 11,932 ton의 담수가 방류되었다(Fig. 2(B)).
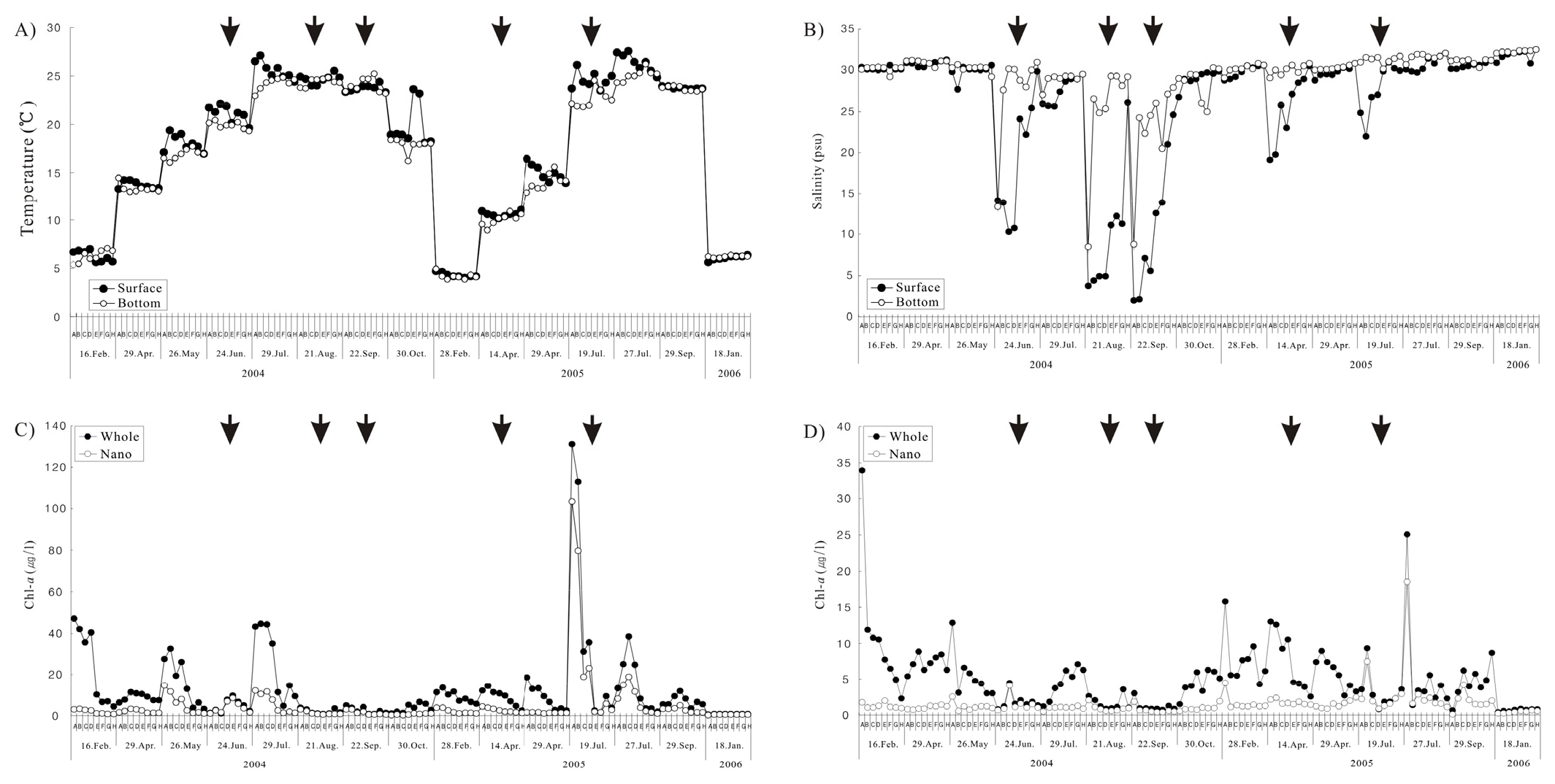
조사기간 동안 수문 개폐에 따른 수온 변화는 개방 시 표층 10.2 (2005년 4월 정점 D)~26.2℃ (2005년 7월 정점 B), 저층 9.0 (2005년 4월 정점 B)~25.2℃ (2004년 9월 정점 F)로 표층과 저층 사이에 차이가 미미하였다. 비개방 시에는 표층 4.0 (2005년 2월 정점 F)~27.6℃ (2005년 7월 정점 C), 저층 3.9 (2005년 2월 정점 F)~26.2℃ (2005년 7월 정점 F)로 표층과 저층의 수온차이는 크게 나타나지 않았으며, 계절에 따른 수온변화는 일반적인 온대해역의 환경을 나타냈다(Fig. 3(A)).
염분의 변화는 수문 개방 시와 비개방 시 현저한 차이를 나타냈다. 개방 시 표층은 2 (2004년 9월 정점 A)~31 (2005년 7월 정점 F), 저층은 8.5 (2004년 8월 정점 A)~31.7 (2005년 7월 정점 H)로 나타나, 전형적인 염쇄기(salt wedge)형 하구의 특징을 보였다. 그러나 비개방 시에는 표층과 저층의 염분 차이가 크지 않았으나, 개방시가 표·저층의 염분은 수문을 열었을 때 낮았다(Fig. 3(B)).
총 엽록소 a농도는 수문 개방 시 표층 0.9 (2004년 8월 정점 E)~131 μgℓ-1 (2005년 7월 정점 A)의 범위로 계절적인 차이가 매우 크게 나타났다. 저층에서는 0.7 (2004년 9월 정점 E)~13 μgℓ-1 (2005년 4월 정점 A)로 수문과 인접한 정점에서 높게 나타났으며, 수문에서 멀어질수록 낮은 농도를 보였다. 비개방 시에는 표층 0.5 (2006년 1월 정점 A)~47.2 μgℓ-1 (2004년 2월 정점 A), 저층 0.4 (2006년 1월 정점 A)~33.9 μgℓ-1 (2004년 2월 정점 A)로 표층과 저층 사이에 큰 차이는 보이지 않았다(Fig. 3(C)). 미소 엽록소 a농도 분포 또한 총 엽록소 a농도 분포와 유사하였으나, 조사 시기 및 정점에 따라 최소 값과 최대 값은 약간 차이를 보였다. 즉 개방 시 표층은 0.5 (2004년 9월 정점 F)~104 μgℓ-1 (2005년 7월 정점 A)를, 저층은 0.4 (2004년 9월 정점 E, F)~7.5 μgℓ-1 (2005년 7월 정점 B)로 나타났다. 비개방 시 표층은 0.3 (2006년 1월 정점 A)~19 μgℓ-1 (2005년 7월 정점 C), 저층은 0.1 (2005년 9월 정점 A)~18.5 μgℓ-1 (2005년 7월 정점 A)로 나타났다(Fig. 3(D)).
3.2 동물플랑크톤의 종조성
동물플랑크톤의 분류군은 수문 개방 시 요각류 21종을 포함하여 총 43분류군이 출현한 반면, 비개방 시에는 요각류 21종을 포함하여 46분류군이 출현하였다(Table 1). 이들 출현 분류군의 공간분포 결과 개방 시와 비개방 시 모두 수문과 인접 정점에서 분류군 수가 높았으며, 수문과 떨어진 정점 H에서 낮은 분류군 수를 보였다.
Table 1.
Opening/closing of gate observation average total abundance (indiv.m-3), number of taxa and dominant species of zooplankton in the study area.
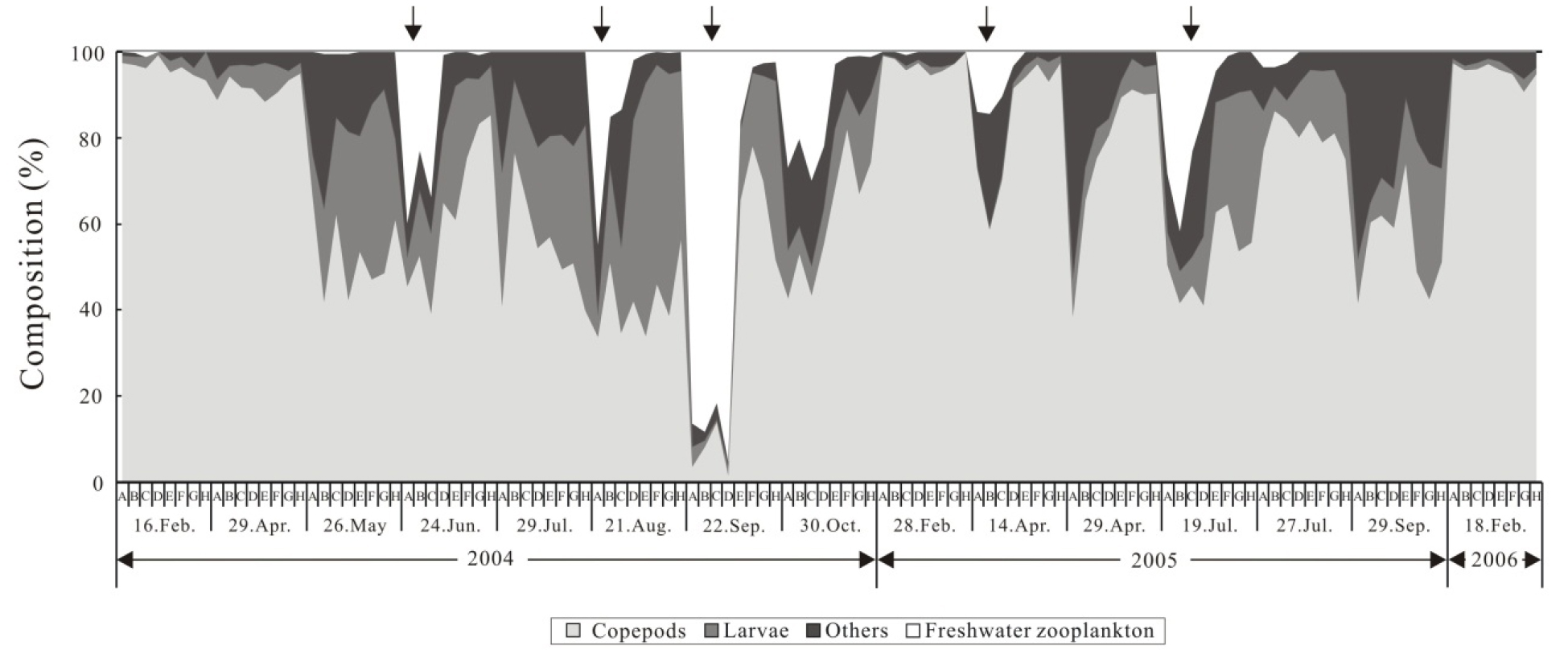
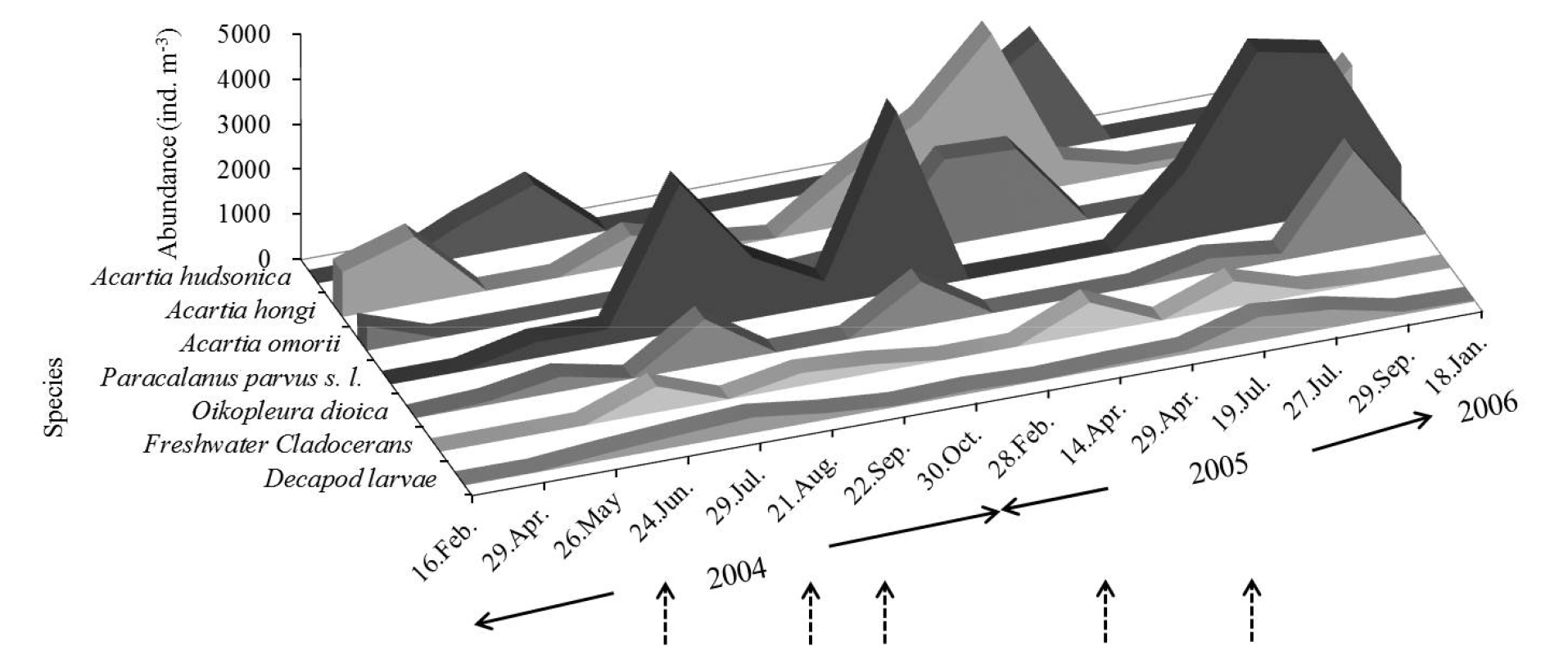
하구역에 출현하는 동물플랑크톤의 주요 분류군을 살펴보면 개방 시 요각류가 높게 나타났으며, 담수 요각류와 담수 지각류, 각 종 동물플랑크톤 유생류, 화살벌레류, 지각류, 미충류 순으로 나타났다. 비개방 시 또한 요각류가 가장 높은 출현을 보였으며, 개방 시 높은 출현을 보인 담수 검물벼룩 요각류와 담수 지각류는 낮은 출현 개체수를 보였다(Fig. 4).
강우의 영향으로 수문이 개방되었을 때, 담수의 급격한 유입으로 인해 동물플랑크톤 출현 개체수는 비개방 시 보다 개방 시에 낮은 출현을 보였으나, 그 차이는 유의하지 않았다(p>0.05) (Fig. 5). 개방 시 출현 개체수는 평균 4,591.2 (287 (2004년 9월 22일 정점 D)~22,843 (2005년 4월 14일 정점 A) ind.m-3) ind.m-3로, 수문 인접 정점인 정점 A에서 높게 출현하였다. 우점 출현한 동물플랑크톤은 Acartia hongi, Paracalanus parvus s. l., A. hudsonica, 담수 지각류, A. omorii, 십각류 유생이었다. 비개방 시 평균 출현 개체수는 5,286.6 (539 (2004년 5월 26일 정점 E)~23,240 (2005년 9월 25일 정점 B) ind.m-3) ind.m-3로, 연안성 요각류인 P. parvus s. l., Acartia hongi, A. hudsonica, A. omorii와 척삭동물인 Oikopleura dioica 순으로 우점하였다(Table 1, Fig. 6).
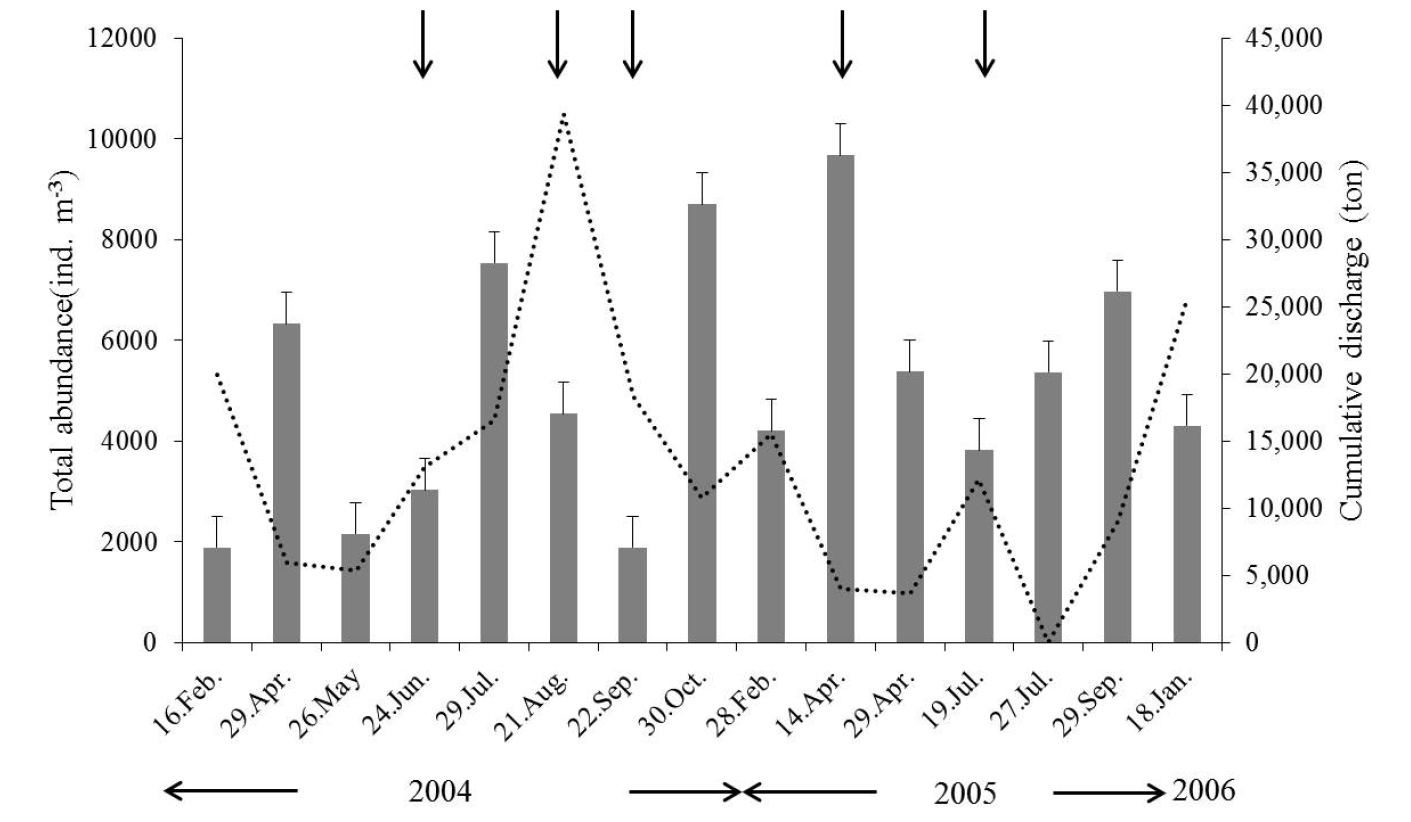
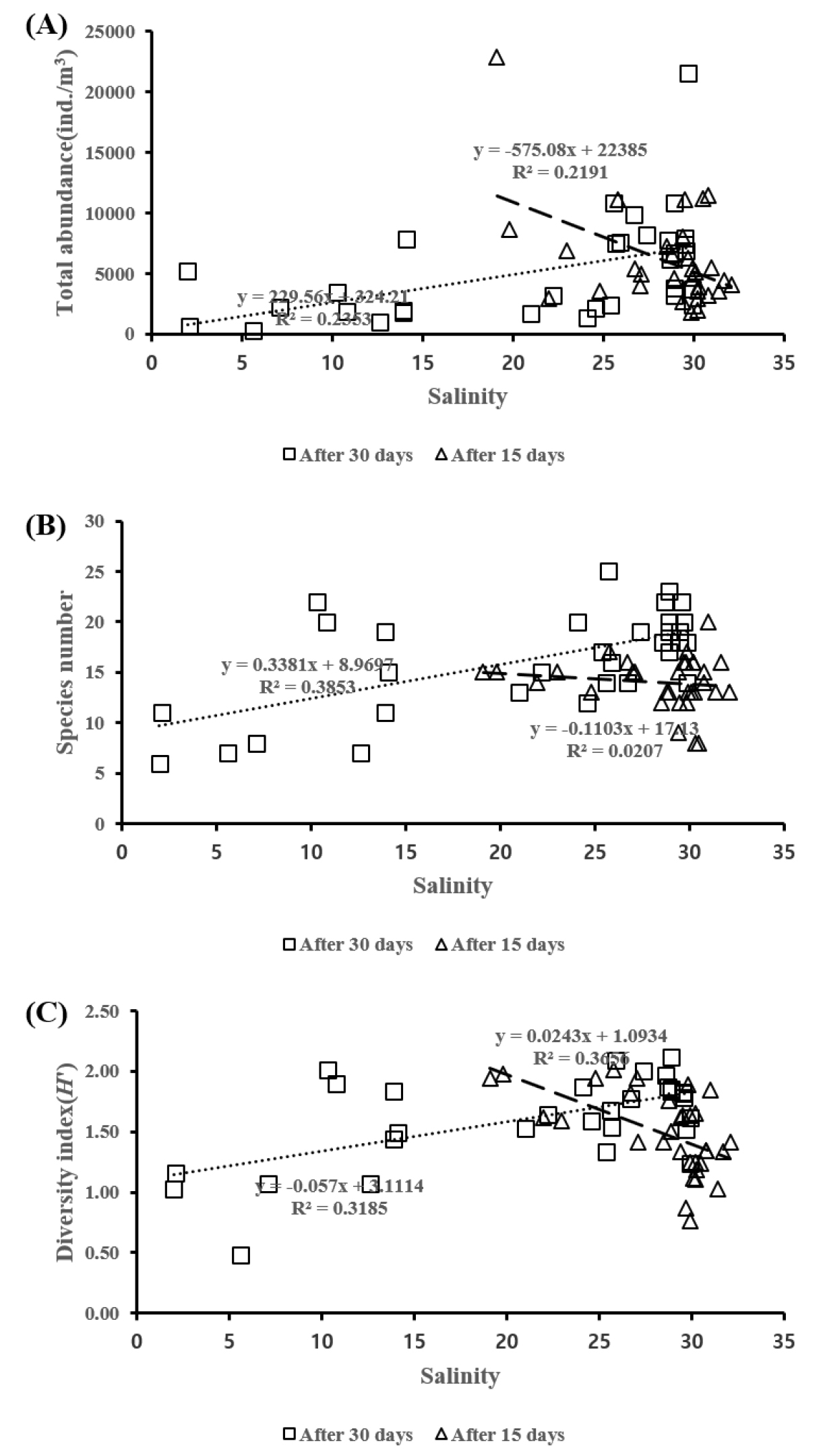
동물플랑크톤의 출현 개체수로부터 계산된 종 다양도 지수는 개방 시 평균 1.17 (2004년 9월 22일)~2.04 (2004년 8월 21일)로 나타났으며, 비개방 시 평균 1.00 (2005년 2월 28일)~1.88 (2004년 7월 29일)로 나타났다(Table 2). 담수의 유입으로 인해 불안정한 환경이 복구되는 시기를 알아보기 위해 수문 개방 후 15일, 30일후 조사를 실시한 결과 15일(p<0.05)의 결과보다 30일 이후(p<0.05) 개체수, 분류군 수, 종 다양성 지수가 높게 나타났다(Fig. 7).
Table 2.
The zooplankton species diversity index appearing in the Yeongan estuary.
3.3 요각류의 출현양상과 주요 우점종
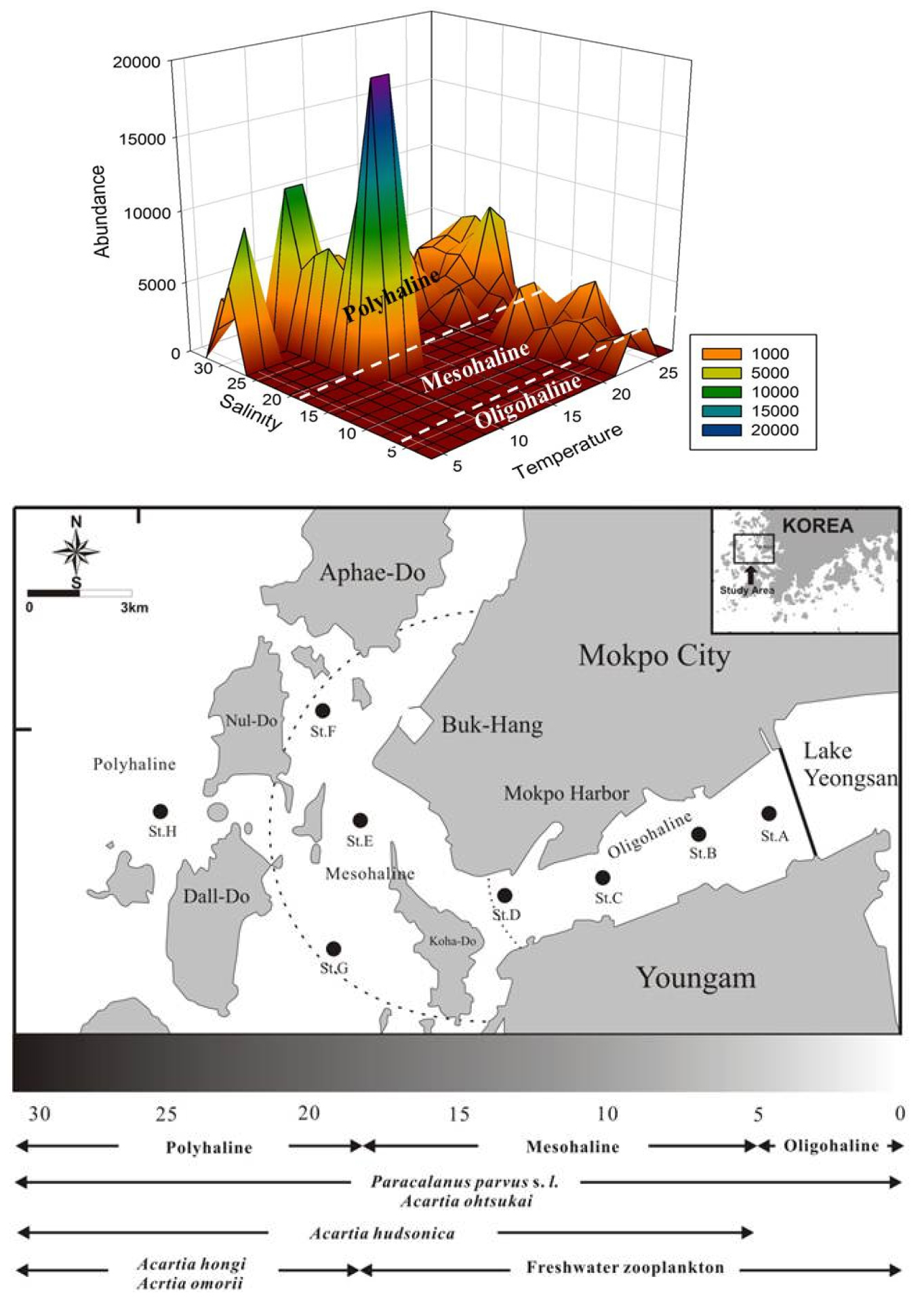
수문 개방 시와 비개방 시 출현한 요각류는 수문 개방 시 담수의 영향을 받은 인접 정점과 연안 정점에서는 담수 요각류의 출현이 높게 나타났으나, 비개방 시에는 연안성 요각류와 기수성 요각류가 우점 출현하였다. 요각류의 출현 개체수도 비개방 시에 높은 출현을 보였다. 개방 시 요각류 출현 개체수는 평균 5,308 ind.m-3로 수문 앞 정점에서 높은 출현개체수를 보였으며, A. hongi, P. parvus s. l., A. hudsonica, A. omorii, 담수 검물벼룩 요각류의 순서로 우점 출현하였다. 비개방 시 평균 출현 개체수는 13,900 ind.m-3로 하구 바깥 정점에서 높게 나타났으며, P. parvus s. l., A. hongi, A. hudsonica, A. omorii, Calanus sinicus가 우점적으로 출현하였다. 강우의 영향으로 하구에 담수의 유입이 매우 큰 8월과 9월의 수문 인접 정점들은 저염분 지역으로 형성되었으며, 중염분 지역은 담수 영향이 적은 수로 내의 정점들로 형성되었다. 고염분 지역은 비개방 시 모든 조사 시기, 수문 인접 정점에서 하구 바깥의 모든 정점으로 나타났으며, 개방 시는 담수의 영향이 적은 장좌도에 위치한 정점들을 포함하였다. 저염분 지역에 출현한 우점 요각류는 검물벼룩 요각류(Cyclopoids)와 담수 지각류, P. parvus s. l., Oithona spp.가 우점 출현하였으며, 중염분 지역에서는 P. parvus s. l., A. hudsonica, 검물벼룩 요각류(Cyclopoids)와 담수 지각류, A. ohtsukai가 우점 출현하였으며, 고염분 지역에서는 A. hongi, A. hudsonica, A. omorii가 우점 출현하였다(Fig. 8).
3.4 우점 출현하는 요각류와 환경과의 상관 관계 및 집괴분석
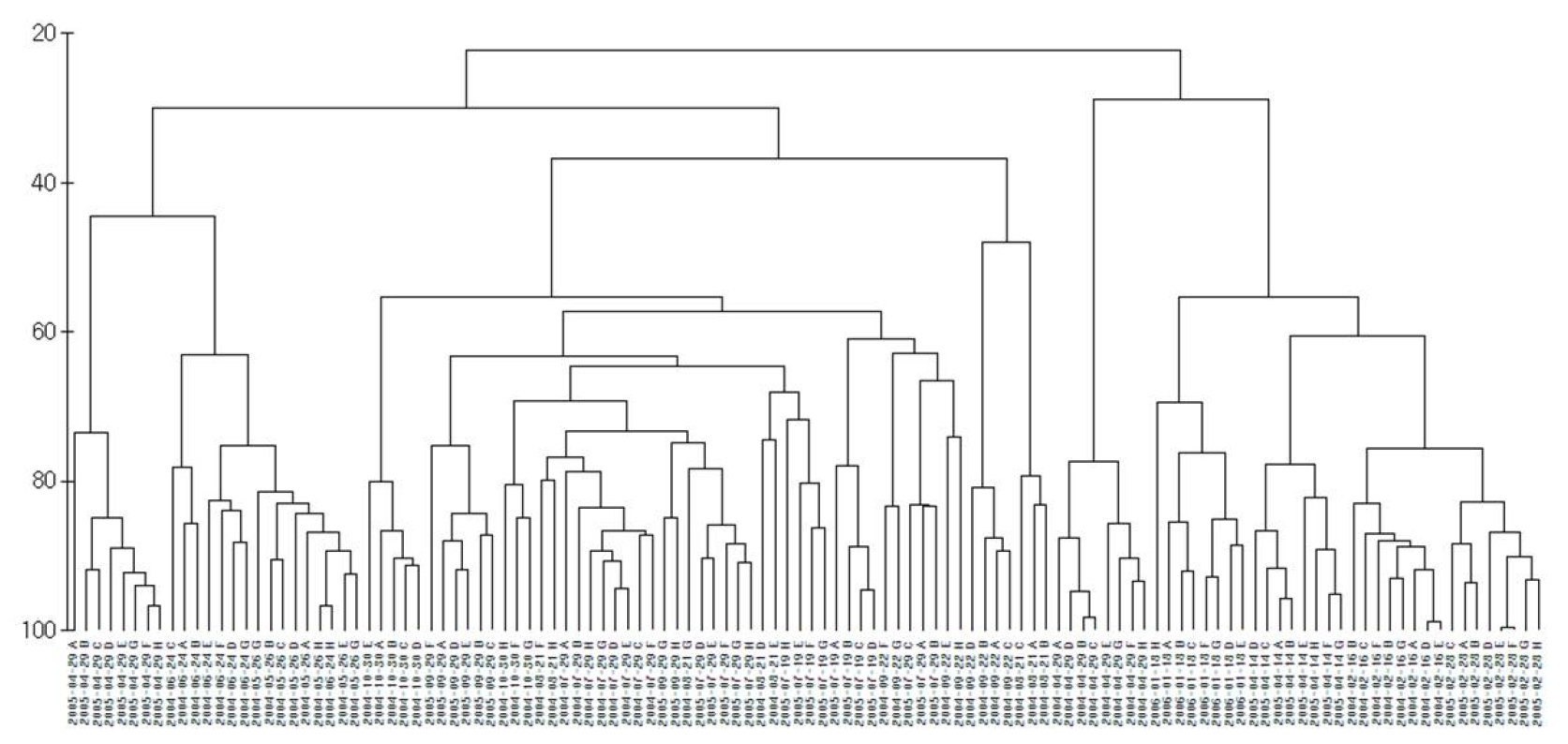
전 연구기간 출현한 요각류의 출현 개체수 자료에 근거하여 집괴분석 결과 개방 시 방조제와 인접한 정점 A~D는 하나의 정점군으로 나타났으며, 담수의 영향을 적게 받는 수로 바깥 정점 E~H는 비개방 시의 정점들과 하나의 정점군으로 나타났다(Fig. 9).
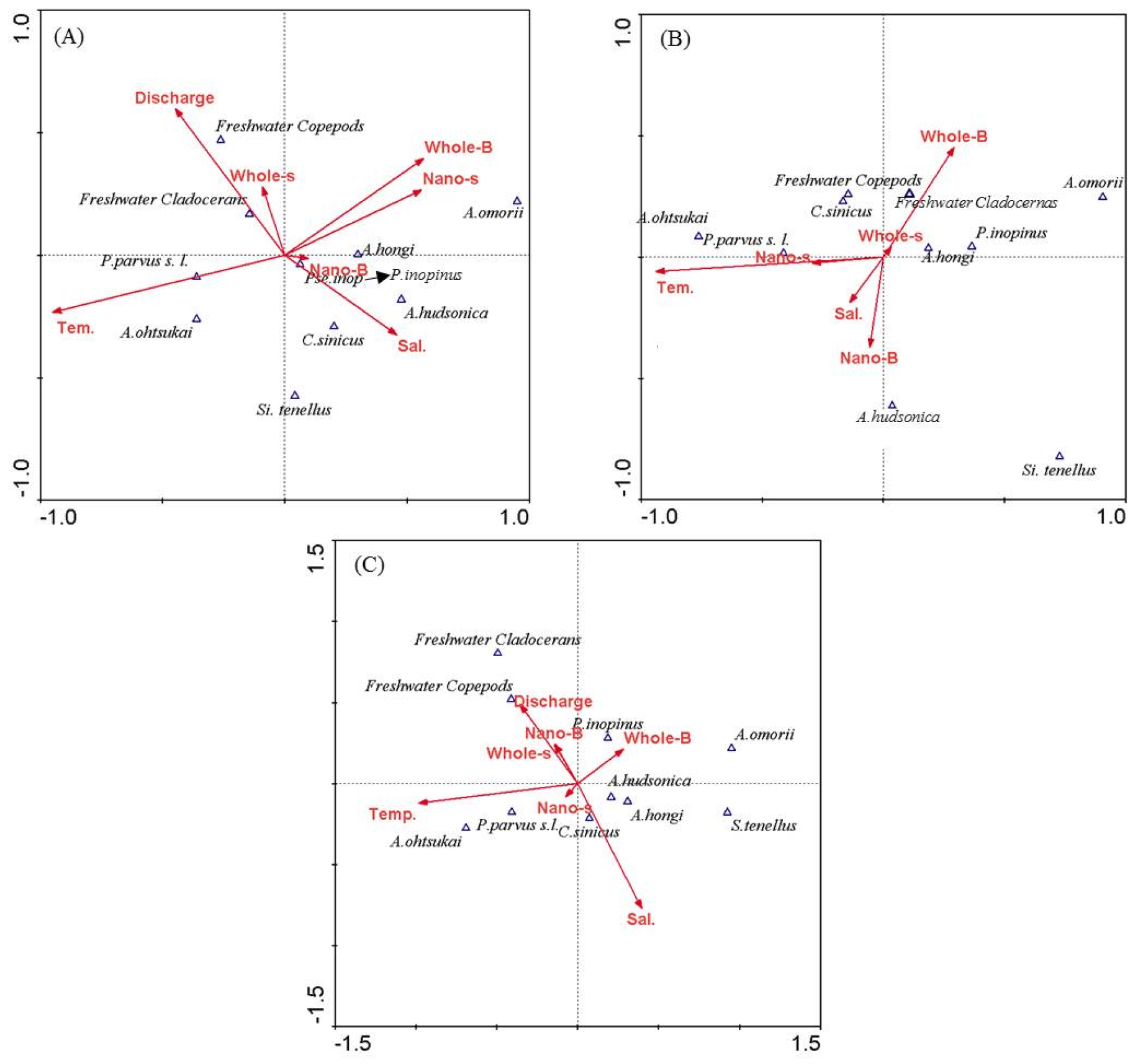
연구 지역에 출현하는 우점 요각류 중 기수성 요각류 및 담수 동물플랑크톤을 대상으로 하여 환경 선호도를 파악하기 위해 수문 개폐 여부에 따라 정준대응분석을 실시하였다. 수문 개방 시 분석결과 제 1축(Axis 1)과 제 2축(Axis 2)의 고유치(eigenvalues)는 각각 0.41과 0.14로 나타났다. 제 1축과 2축의 전체 자료에 대한 누적 기여율은 51.4%로 나타났으며, 제 1, 2축에서 환경요인과 동물플랑크톤의 변화에 대해 77.1%를 설명하였다(Table 2). 그 결과 P. parvus s. l., A. ohtsukai는 수온과 염분에 양의 상관성을 보였으며, A. hongi, A. hudsoinica, A. omorii, Pseudodiaptomus inopinus는 수온에 음의 상관성을, 염분에 양의 상관성을 나타냈다. 담수동물플랑크톤은 방류량과 양의 상관성을, 염분에 음의 상관성을 나타냈으며, Calanus sinicus는 염분에 양의 상관성을 나타냈다(Fig. 10(A)).
비개방 시 분석결과에서 제 1축과 제 2축의 고유치는 각각 0.21와 0.07의 고유치를 보였다. 제 1축과 2축의 전체 자료에 대한 누적 기여율은 18.5% 으며, 제 1, 2축에서 환경요인이 동물플랑크톤의 변화에 대해 82.5%를 설명하는 것으로 나타났다(Table 3). 그 결과 Calanus sinicus는 염분과 양의 상관성을 나타냈으며, 나머지 종들은 비개방 시의 결과와 동일 하였다(Fig. 10(B)).
Table 3.
Summary of Canonical Correspondence Analysis (CCA) for dominant copepods abundance in the Yeongan estuary.
4. 고찰
영산강 하구는 하구둑과 금호 방조제가 건설된 후 인접 해역과 해수 교환이 원활하게 이루어지지 않고 있으며, 해양 쪽으로 나가는 흐름만 존재하고 유입은 거의 없어 하구의 순환이 조석에 의해 큰 영향을 받고 있다(Kang 1998). 그러나 하계에는 댐의 수위 조절로 인한 수문의 개방으로 오염된 영산호의 담수가 일시에 유입되어, 폭이 좁은 목포 수로를 통해 외해와의 해수 교환이 일어나고 있다(Park et al. 2001).
동물플랑크톤의 먹이량을 가늠해 볼 수 있는 식물플랑크톤 엽록소 a농도는 수문 개방 시 담수 유입으로 인한 환경의 불안정한 현상으로 낮은 농도를 나타냈고, 비개방 시에는 환경의 안정화로 높은 농도를 보였다. 이는 영산강 하구역의 짧은 항내 거리와 일시적 담수 유입에 의한 유속(Flushing rate) 증가가 엽록소 a농도 변화에 영향을 미친 것으로 보인다. 엽록소 a농도는 수문 개방 시 다량의 담수와 함께 유입되는 부유물의 증가로 투명도가 크게 감소하며, 식물플랑크톤이 이용할 수 있는 빛의 양이 제한됨으로 직접적인 영향을 받는다(Jeong 2008). 높은 영양염이 존재하는 tidal freshwater지역에서 유속의 영향과 부유물질 유입에 따른 탁도의 증가는 엽록소 a농도 및 식물플랑크톤 생산력에 큰 영향을 미친다(Arruda et al. 1983; Hart 1988; Kirk 1991; Sin et al. 2005). 또한 하구역에서 염분 변화는 동물플랑크톤 군집구조를 결정하는 중요 요인 중 하나이다(Heydorn and Grindley 1981; Jerling 2003). 생물은 삼투 조절이라는 면에서 염분 변화에 민감하며, 수서 생물의 경우 이동 능력이 있어 적정한 염분 범위를 찾아 서식지를 선택할 수 있으나, 동물플랑크톤은 이동능력이 매우 약해 서식지의 염분 변화에 따라 크게 좌우된다(Gunter 1961; Jones et al. 1990). 영산강 하구는 수문 개방 시 하구 안쪽의 표층 염분은 비개방 시에 비해 최대 표층 30.5, 저층 20.0의 농도 차이를 나타냈다. 담수 유입에 의한 염분 변화는 하구 내 정점에서 급격히 변동하였으며, 외달도에 위치한 정점 H까지 미미한 영향을 나타냈다. 영산강 하구내 염분 농도 변화는 건기와 우기 사이에 큰 차이를 보였으며, 육상에서 유입된 담수 검물벼룩 요각류(Cyclopoids)와 담수 지각류는 하구 입구까지 나타났다. 하구내 담수 유입으로 염분의 변동은 동물플랑크톤의 출현 개체수, 종조성 및 분류군수에 영향을 미치나, 하구 내 일부 요각류는 이러한 급격한 환경변화에 적응할 수 있다(Cronin et al. 1962; Jeffries 1962; Knatz 1978; Lee and McAlice 1979; Durbin and Durbin 1981; Fulton 1984). 비개방시 P. parvus s. l., A. hongi, A. hudsonica, A. omorii, A. ohtsukai, Eurytemora pacifica가 우점 출현하였으며, 개방 시 담수 검물벼룩 요각류와 담수 지각류(Cladocerans)가 함께 우점 출현하였다. 그러나, 수문을 개방하면 비개방시에 우점하였던 Oikopleurda dioica와 따개비 유생은 사라지고, 복족류 유생, 십각류 유생 및 해파리류(Unidentified Trachymedusae)가 우점 출현하였다. 하구둑 수문 개폐 여부에 상관없이 염분 범위만을 고려하여 하구를 구분한 결과, 저염분 지역은 연안성 동물플랑크톤의 개체수 감소와 담수 동물플랑크톤의 대량 유입이 나타났으며, 중염분 및 고염분 지역으로 갈수록 담수 동물플랑크톤의 개체수 감소와 연안성 동물플랑크톤 개체수가 증가하였다. 새만금 방조제 설치 이전 만경∙동진강 하구의 동물플랑크톤 군집은 연안성 동물플랑크톤으로 구성되었으며, 넓은 범위의 염분에서 서식이 가능한 광염성 종들이 출현하였다(Suh et al. 1991). 그러나 새만금 방조제 건설 후 여름철 집중호우로 방조제 전 지역에 저염분의 확산으로 담수종 및 기수종의 증가를 예상하였으나 광염종인 P. parvus s. l.가 높게 나타나, 새만금 해역의 동물플랑크톤 분포는 인위적인 방 조제의 수문개폐로 인한 해수의 유입량에 따라 결정될 수 있다는 것을 뒷받침해준다(Lee et al. 2009). Cochin 하구는 수온 변동은 적으며, 우기에 접어들면 염분 변화가 매우 심각하게 나타나 약 30에서 담수에 가깝게 희석되며, 하구역에 출현하는 Acartia속 9종은 계절(수온)과 염분 변화에 따라 천이 양상이 결정되며, 특히 Acartia centrura와 A. spinicauda, A. bilobata와 A. southwelli는 염분 변화와 종간 경쟁에 민감하게 반응한다(Tranter and Abraham 1971). Siene 하구에 출현하는 Temora longocirnois, Centropages spp., Oikopleura dioica가 고염분 지역에서 출현한 반면, 중염분~저염분 지역은 Acartia spp.가, 저염분 지역은 담수 요각류인 Acantocyclops robutus, Diaptomus spp., 담수 지각류가 출현하였다. 고염분 지역에서 저염분 지역으로 갈수록 광염성 동물플랑크톤의 개체수는 감소를 한 반면, 담수 동물플랑크톤의 개체수가 증가하였다(Mouny and Dauvin 2002). 인위적으로 수문이 개방이 되었을 때 영산강 하구 환경은 개방형 하구 형태인 섬진강 하구의 풍수기 환경과 같이 많은 양의 담수 유입과 하구 내 염분 농도 감소로 담수 동물플랑크톤이 하구내에 출현하였다. 하구둑이 건설되지 않은 섬진강 하구는 강우 후 동물플랑크톤의 분류군 수는 증가한 반면, 출현 개체수는 감소하였다. 이는 짧은 시간 많은 양의 담수가 하구 내 유입으로 담수 동물플랑크톤의 가입과 안정적인 염분 환경을 불안정하게 변동시켜 하구에 서식하는 동물플랑크톤 군집까지 큰 영향을 미쳤다(Kim et al. 2000). 영산강 하구는 수문 폐쇄로 담수 공급이 차단되면, 하구 내 해수의 유입으로 염분 농도가 서서히 회복됨에 따라 동물플랑크톤이 연안 광염종으로 우점종이 바뀌며, 출현 개체수 또한 증가하였다. 수문 개방으로 인한 담수의 유입이 염분 농도의 변화를 유발하여 단기적으로는 식물플랑크톤의 생체량 감소를 초래하나, 영얌염류 등의 공급으로 장기적으로는 식물플랑크톤 생체량이 증가하게 되며, 이는 일차소비자인 동물플랑크톤 현존량 증가에 긍정적으로 작용하는 것으로 판단된다. 그러나, 이러한 긍정적인 효과가 일시개방형 하구에서 어느 정도 지속될 수 있는지에 대해서는 보다 짧은 채집 간격에 의한 동물플랑크톤 동태 연구가 필요할 것으로 사료된다.
