

1. 서론
2. 재료 및 방법
2.1 조사지 개황
2.2 시료 채집 및 수질환경 측정
2.3 식물플랑크톤 정량 및 정성 분석
3. 결과 및 고찰
3.1 식물플랑크톤 모니터링
3.2 주요 적조 원인종의 형태학적 특징
3.3 겨울철 석호의 식물플랑크톤 대발생에 대한 첫 보고
1. 서 론
석호는 해안선을 따라 분포하는 하천의 하구가 연안류의 작용 등으로 생성된 사주 또는 사취의 모래톱에 의해 입구가 막힘으로써 바다와 격리되어 생성된 자연호수이다. 국내에서 석호는 동해안 일대에 112 km의 해안선을 따라 조석의 영향이 적은 곳에 18개가 분포한다. 동해안의 석호는 최종빙기 최성기의 해수면 하강과 해진극상기의 해수면 상승을 거듭하며 연안류의 작용에 의한 침식과 퇴적을 반복하여 6,000년경 생성된 이후 현재까지 유지되어 왔다(Yoon et al. 2008). 생태학적으로 내륙에서 유입되는 담수와 해수가 공존하는 기수호로 담수와 해수생태계의 중간단계인 전이생태계로써 특징에 의해 담수, 기수 및 해양생물이 공존하여 생물다양성이 매우 높다(Basset et al. 2013a; Basset et al. 2013b). 또한 자연적으로 형성된 수려한 주변환경으로 다양한 볼거리와 함께 심미적 휴식과 여가선용 공간을 제공하여 관광자원으로써 활용가치가 높다. 이와 같이 석호는 지질학적, 생태학적 및 관광자원으로써 높은 가치가 있음에도 불구하고, 매립과 난개발로 그 규모와 경관보존율이 크게 축소되거나 변해왔다(Yoon et al. 2008).
동해안의 대표적인 석호 중의 하나인 영랑호는 하상구조가 주로 모래와 뻘로 이루어져 있으며, 주변은 콘도, 순환도로, 공원이 조성되어 있어 인간의 유입이 용이하다. 영랑호 하구의 입구는 두텁게 형성된 사주에 의해 폭이 좁고, 도로 및 건물 등이 건설되어 있어 해양생태계와 연결(갯터짐)이 정상적으로 발생하지 않아 석호로서의 기능이 저하되었다(Lee et al. 2006). 영랑호에 대한 과학적 연구는 환경 및 플랑크톤의 관계(Cho and Park, 1969)를 시작으로 수질평가 연구(Huh et al. 2017; Lee and Kwak 1987), 지형특성 및 퇴적환경 연구(Lee et al. 2006; Yoon et al. 2008) 및 고기후 연구(Nakai and Hong 1982) 등의 연구가 수행되어 왔으며 대부분이 관광과 개발로 인한 수질상태 및 경관 변화 등에 집중되어 왔다. 자연석호의 특성상 담수와 해수생태계의 전이대로써 높은 생물다양성을 가지는 잠재성에도 불구하고 영랑호에서 생물상에 관한 연구는 일부 어류상 연구(Choi et al. 2007)와 최근 영랑호를 포함한 동해안 4개 석호를 대상으로 한 식물플랑크톤 연구(Moon et al. 2015)를 제외하고 전무한 실정이다.
적조는 수서생태계에서 플랑크톤의 이상 증식에 의해 바다, 강, 호소 등이 변색되는 현상으로 경제적, 생태학적, 심미적 문제를 야기한다. 국내 적조 발생은 연안성 와편모조류, 침편모조류 또는 규조류 등이 남해 연안에서 주로 발생되어 왔다(Yoon 2011). 석호와 같은 고립된 해양 지역에서 적조 발생에 관한 기록은 전세계적으로 드물게 보고되어 왔으나(e.g. (Daghor et al. 2018; Maclean 1975; Morton and Villareal 1998; Robinson and Brown 1983), 국내 석호에서 적조발생에 관한 기록은 아직까지 보고된 바 없다. 본 연구는 2009년 1월, 겨울철 영랑호에서 적조 발생에 관한 첫 보고와 함께 적조 원인종을 규명하고, 영랑호의 식물 플랑크톤 대발생 양상을 확인하고 위하여 2009년 1월부터 2010년 4월까지 조사를 수행하였다. 본 연구는 비록 과거의 연구결과이나 석호에서 어떤 식물플랑크톤이 대발생 하는가에 대한 정보 제공이 학술 논문의 가치가 있다고 판단하여 보고하는 바이다.
2. 재료 및 방법
2.1 조사지 개황
영랑호는 강원도 속초에 위치한 자연석호로 면적은 2008년 기준 0.96 km2이고, 호 안 길이는 7.21 km로 동해안 14개 석호 중 규모가 두번째로 크다. 20세기 초부터 1990년대까지 그 규모가 지속적으로 감소해왔으나, 그 이후 인공호수로써 호 안이 정비되면서 그 면적은 유지되고 있다(Yoon et al. 2008). 영랑호의 상류인 서쪽 부분은 담수의 주요 유입원인 장천과 연결되고, 하류인 동쪽 부분은 지속적인 해수의 주요 유입원인 동해와 연결된다(Fig. 1). 바다와 연결되는 하류는 하나의 좁은 물길에 의해 동해와 이어지나 모래톱에 의해 빈번히 막혀 해수와의 교류가 종종 제한되어 연중 염분 수준은 20 psu를 넘지 않는다(e.g. Moon et al. 2015).
2.2 시료 채집 및 수질환경 측정
식물플랑크톤 채집과 수질환경 측정은 2009년 1월 23일 영랑호 식물플랑크톤 대발생을 최초 관찰한 이후, 2009년 2월 및 2010년 4월까지 총 5회(2회는 영랑호 결빙에 따라 조사 불가능)에 걸쳐 영랑호 3개 정점에서 수행되었다(Table 1, Fig. 1). 수온, 염분 및 용존산소는 휴대용 Portable T-S meter (YSI, model 85, USA)로 현장에서 측정하였고, pH는 실험실에서 채수된 시료 일부로부터 pH 미터기(Thermo Scientific, Orion 2-Star, USA)로 측정하였다. 식물플랑크톤의 정량 및 정성분석을 위한 시료채집은 시수 1 L를 폴리에틸렌 채수병에 담아 Gultaraldehyde 고정액을 사용하여 최종농도 2% 로 고정한 후 실험실로 운반하였고, 침전법에 의해 150~200 ml로 농축하여 공시재료로 하였다.
Table 1.
Sampling information in the Young Rang lagoon (ND: No Data).
2.3 식물플랑크톤 정량 및 정성 분석
식물플랑크톤의 정량분석은 농축된 시료를 희석병에서 균일하게 희석한 후 1 ml를 취하여 Sedgwick-Rafter 챔버에 넣고 광학현미경(Zeiss, Axioskope 40, German) 400배하에서 3회 계수한 후 평균치를 단위체적당 개체수로 환산하였다. 식물플랑크톤의 정성분석은 광학현미경 1,000배하에 관찰하였고, 주요 우점종의 영양형을 구별하기 위해 Primulin 염색 후에 형광현미경하에 광합성을 위한 색소체 유무를 확인하였다(Yamaguchi et al. 1995). 광학현미경하에 동정이 어려운 종에 대해, 미세구조를 관찰하기 위하여 Jung et al. (2010)의 방법에 따라 주사전자현미경(Jeol, JSM 5600LV, Japan)으로 관찰하였다.
3. 결과 및 고찰
3.1 식물플랑크톤 모니터링
조사기간 중 식물플랑크톤 개체수는 2009년 1월 23일 적조 발생시 평균 4.68±0.40×1010 cells mL-1를 보였으며, 3주 후인 2월 17일 조사시 3.82±1.40×104 cells mL-1로, 기존 발생되었던 식물플랑크톤 대발생은 소멸하였다. 2009년 1월 대발생 이후, 2010년도 재발생을 확인하기 위하여 저수온기(1–4월)까지 5회 조사를 수행하였으나, 1–2월(2회 조사)의 영랑호 결빙을 제외하고 평균 식물플랑크톤 개체수는 1.07±0.58×104 cells mL-1로 적조가 발생되지 않았다. 영랑호에서 겨울철 대발생은 주기적으로 발생하는 현상이 아닌 일시적인 현상에 의한 것으로 판단된다.
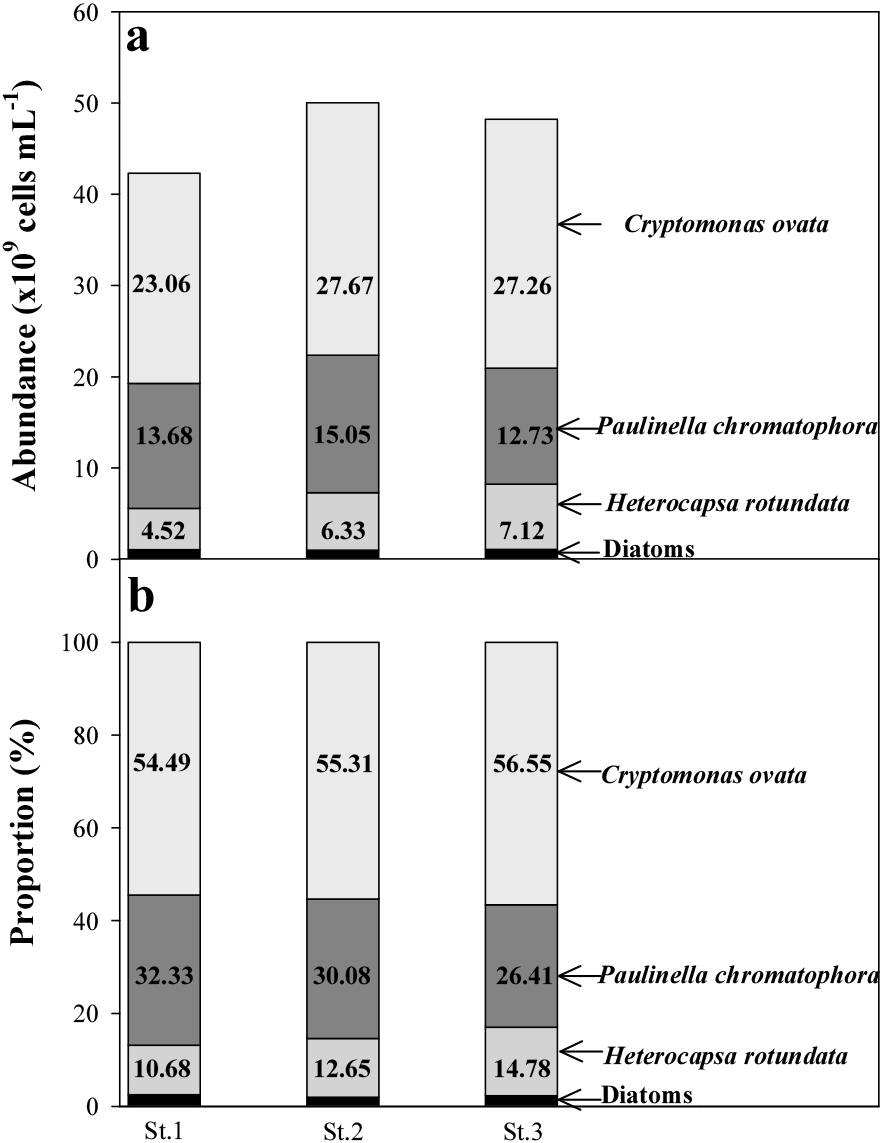
2009년 1월 23일 영랑호 조사 당시 호수색은 식물플랑크톤 대발생으로 인하여 붉은색을 띄었고(Figs. 2A & B), 식물플랑크톤 3종[녹조류 Cryptomonas cf. marssonii Skuja, 아메바류 Paulinella ovalis (Wulff) Johnson, Hargraves & Sieburth, 와편모조류 Heterocapsa rotundata (Lohmann) G. Hansen]이 압도적으로 우점하였다(Figs 2C, D). 즉, C. cf. marssonii는 2.60±0.26× 1010 cells mL-1 개체수로 55.5%를 차지하여 제1우점하였고, P. ovalis는 1.38±0.12×1010 cells mL-1 개체수로 29.6%를 차지하여 제2우점하였으며, H. rotundata는 5.99±1.33×109 cells mL-1의 개체수로 12.7%를 차지하여 제3우점하여, 3종이 총 개체수 대비 97.76%를 차지하였다(Fig. 3).
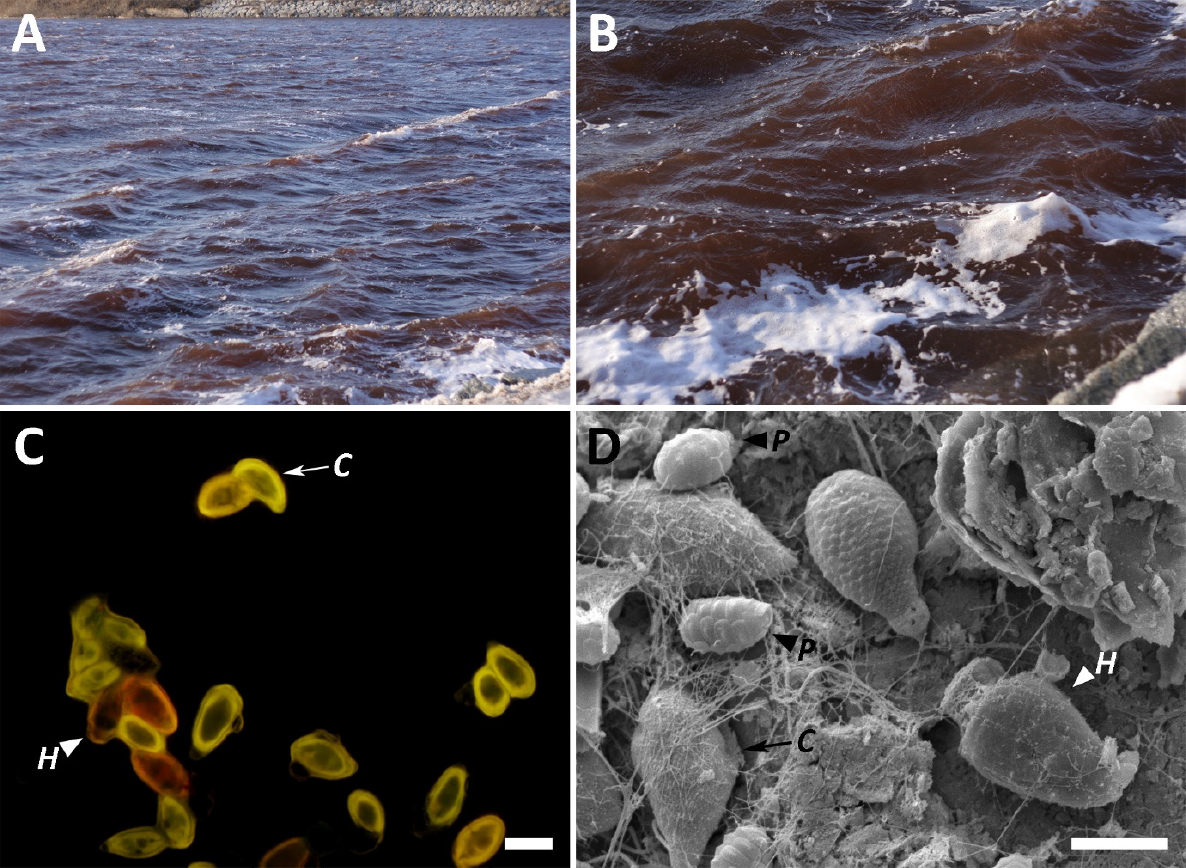
Fig. 2
Red-tides in Youngrang lagoon (A & B). Cryptomonas cf. marsonii (arrow) and Heterocapsa rotundata (white arrowhead) in fluorescence microscope (C). Three red-tides caused species, Cryptomonas cf. marsonii (arrows), Paulinella ovalis (black arrowhead) and Heterocapsa rotundata (white arrowhead) in scanning electron microscope (D).
3.2 주요 적조 원인종의 형태학적 특징
KINGDOM Chromista Cavalier-Smith, 1981
PHYLUM Cryptophyta Cavalier-Smith, 1986
CLASS Cryptophyceae Fritsch, 1937
ORDER Cryptomonadales Pascher, 1913
FAMILY Cryptomonadaceae Ehrenberg, 1831
GENUS Cryptomonas Ehrenberg, 1831
Cryptomonas cf. marssonii Skuja, 1948 (Figs 4A & B)
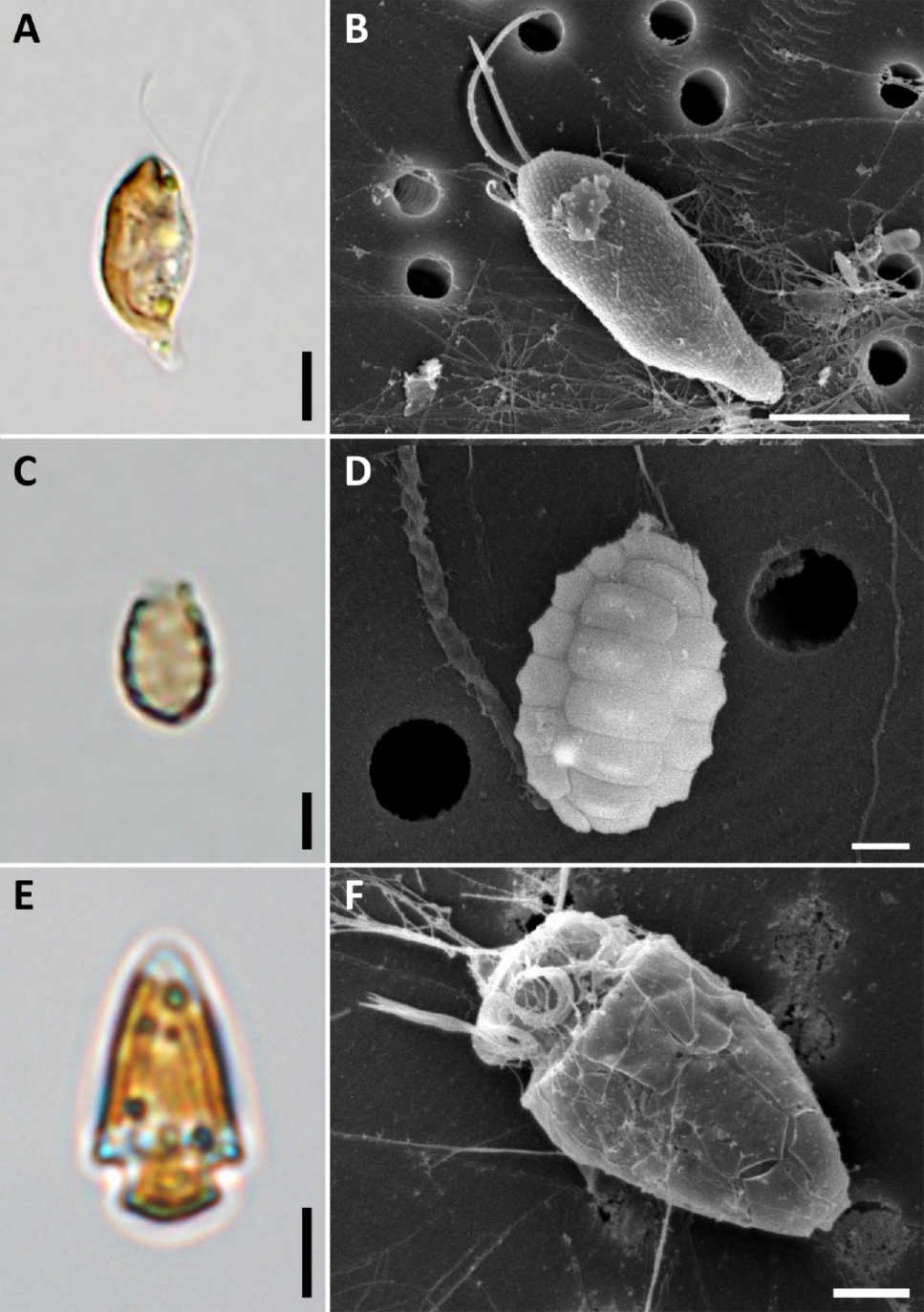
형태학적 특징: 세포는 S자형으로 휘어지고, 배면 보기에서 비대칭하다(Figs 4A & B). 세포 길이는 9.9–13.1 μm이고, 세포 끝은 antapex에서 부리형으로 좁아진다. 광학현미경에서 엽록체는 세포의 가장자리에 위치하고 중앙에 피레노이드가 있으며, 그 사이에 전분 알갱이가 분포한다(Fig. 4A). 주사전자현미경에서 다수의 비늘형 돌기가 세포표면을 덮고 있다(Fig. 4B).
생태학적 특징:Crpytomoans marssonii는 스웨덴 웁살라의 담수의 호수로부터 최초로 기재되었고(Skuja 1948), 이후 전세계 담수 지역에서 빈번히 보고되어 왔으며(e.g. (Javornický 2014; Kim et al. 2007; Xia et al. 2015), 일부 지역에서는 대발생을 야기한 바 있다(Kobanova et al. 2016; Pithart 1997). 국내에서 C. marssonii는 나주의 오정 연못과 함안의 대평늪, 가항늪, 질랄늪 등의 담수에서 처음으로 보고되었다(Kim et al. 2007). 담수와 해수의 전이지역인 석호와 같은 기수역에서 일부 Crytomonas 종들은 기록된 바 있으나, C. marssonii의 기록은 보고된 바 없다. 본 연구동안 영랑호에서 가장 우점한 Cryptomonas 종은 형태적으로 S자형 세포와 세포 말단에서 부리형으로 좁아지는 특징에 의해 C. marssonii와 유사하나, 생태학적 분포에서 담수에서만 보고된 C. marssonii와 다르다. 기수호인 영랑호에서 Crpytomonas의 추가적인 연구가 보완될 때까지 본 연구동안 적조를 야기한 Cryptomonas 종을 C. cf. marssonii로써 보고한다.
PHYLUM Cercozoa Cavalier-Smith, 1998
CLASS Imbricatea Cavalier-Smith, 2003
ORDER Euglyphida Copeland, 1956 emend. Cavalier-Smith, 1997
FAMILY Paulinellidae De Saedeleer, 1934
GENUS Paulinella Lauterborn, 1895
Paulinella ovalis (Wulff) Johnson, Hargraves & Sieburth, 1988 (Figs 4C and D)
기본명:Calycomonas ovalis Wulff, 1919
형태학적 특징: 껍질은 길게 신장된 아원통형이며(Fig. 4C), 길이는 4.5–5.7 µm이고, 너비는 2.9–3.4 µm이다. 규산질의 인편은 가로로 넓은 직사각형에서 육각형이며, 각 비늘은 6–7개 열로 겹치지 않게 길이방향으로 배열하여 하나의 열을 이루고, 총 5개의 인편열이 하나의 세포를 구성한다(Fig. 4D). 상단부는 3개의 넓은 비늘이 타원형의 헛소공을 구성한다. 광합성 색소체인 cyanelle은 관찰되지 않는다.
생태학적 특징:Paulinella ovalis는 전세계적으로 분포하고(Johnson et al. 1988), 일부 지역에서 늦은 봄에서 여름 사이에 종종 대발생을 일으켰다: 미국 노스캐롤라이나의 팜리코 강(Stanley and Daniel 1985)과 미국 로드아일랜드 나라간셋 만(Johnson et al. 1988).
PHYLUM Miozoa Cavalier-Smith, 1987
CLASS Dinophyceae Fritsch, 1927
ORDER Peridiniales Haeckel
FAMILY Heterocapsaceae R.A.Fensome, F.J.R.Taylor, G.Norris, Sarjeant, Wharton & G.L.Williams, 1993
GENUS Heterocapsa Stein, 1883
Heterocapsa rotundata (Lohmann) G. Hansen, 1995 (Figs 4E & F)
기본명:Amphidinium rotundatum Lohmann, 1908
형태학적 특징: 세포는 총알 모양이고, 상각은 하각에 비해 더 크다(Figs 4E & F). 길이는 9.8–14.8 μm이고, 너비는 5.1–8.1 μm이며, 덮개판은 Po, cp, 5′, 3a, 7′′, 6c, 5s, 5′′′, 2′′′′로 배열한다. 핵은 구형으로 하각에 위치하고, 피레노이드는 역시 구형이며 전분막에 둘러싸여 있고 상각의 하단에 위치한다. 엽록체는 하나이고, 황갈색이다.
생태학적 특징:Heterocapsa rotundata는 범세계적 종으로서 온대역과 아열대역의 연안에서 보편적으로 출현한다고 알려져 있으며(Steidinger and Jangen 1997), 우리나라의 경우 시화호 및 새만금호 등과 같은 폐쇄성 수역에서 적조를 야기한 바 있다(Seong et al. 2006).
3.3 겨울철 석호의 식물플랑크톤 대발생에 대한 첫 보고
국내 식물플랑크톤 대발생에 관한 과학적 기록은 1961년 10월 진해만에서 규조류 Chaetoceros속과 Nitzschia속의 대발생에 관한 보고를 시작으로(Yoon 2011), 1980년대부터 최근까지 남해안에서 늦여름에서 가을철에 주로 기록되어 왔으나(Yoon 2011), 겨울철 동해안 기수역인 석호에서 대발생에 관한 기록은 전무하다. 본 연구는 동해안 대표 석호인 영랑호에서 겨울철에 발생한 식물플랑크톤 대발생에 관한 첫 보고이다. “하구, 석호 육해전이수역 통합환경관리 방안 연구 보고서”에 따르면, 영랑호의 수질은 부영양 상태를 넘어 과영양 상태를 나타내었고, 이는 관광지와 주변 아파트의 하수에 의한 수질악화가 주원인이라 보고하였다(Lee et al. 2001). 또한 2000년에는 수환경 개선을 위한 퇴적물 준설이 있었고(Huh et al. 2017), 2008년 8월을 전·후로 태풍에 의한 호수 수체의 전도로 저층 황화물의 확산에 의해 호수 내 어류들이 폐사한 바 있다(Kim et al. 2008). 이전 영랑호 환경에 관한 보고서와 학술발표자료에 근거하여, 본 조사가 수행된 2009년 겨울철 당시 영랑호는 부영양화가 상당히 진행된 상황이라 추정되며, 이는 겨울철 식물플랑크톤 대발생의 주된 원인이 되었을 것으로 판단된다.
국립수산과학원과 지역자치단체의 적조모니터링의 누적된 자료를 바탕으로 국내 적조현황에 대해 정리한 결과에 따르면, 1972년 적조 모니터링이 시작된 이후 적조 원인종은 시기에 따라 변해왔다. 즉, 1970년대 적조발생은 Skeletonema costatum (Greville) Cleve, Chaetoceros spp., Nitzschia spp., Leptocylindrus danicus Cleve와 같은 규조류에 의해 주로 발생하였고, 1980년대에는 Karenia mikimotoi (Miyake & Kominami ex Oda) Gert Hansen & Ø.Moestrup, Prorocentrum spp., Heterosigma akashiwo (Y. Hada) Y. Hada ex Y. Hara & M. Chihara등 연안성 와편모조류 및 침편모조류에 의해 발생하였으며, 1993년 이후부터 최근까지는 와편모조류인 Margalefidinium polykrikoides (Margalef) F.Gómez, Richlen & D.M.Anderson (=Cochlodinium polykrikoides Margalef)가 적조현상의 주요 생물군이었다(Lim et al. 2020). 2009년 겨울철 영랑호에서 주로 우점한 3종의 식물플랑크톤, Cryptomonas cf. marssonii, Paulinellao ovalis, Heterocapsa rotundata에 의한 혼합 적조 발생은 이번 연구에서 처음으로 보고된다. Konoike et al. (2015)은 부영양화가 진행된 바이칼 호수에서 Cryptomonas marssonii의 대발생을 보고하였다. Xia et al. (2015)은 Cryptomonads의 개체수가 pH, 용존 질소 및 용존 탄소와 양의 상관관계에 있음을 확인하고, 수계 내 영양화 수준이 개체수에 영향을 미치는 결정적 요인이라 제안하였다. 겨울철 영랑호에서 Cryptomonas cf. marssonii의 대발생은 부영양화에 의한 것이라 판단된다. Johnson et al. (1988)은 1985년 여름 나라간셋 만에서 Aureococcus anophagefferens과 함께 P. ovalis의 혼합 대발생을 발견하였고, 1986년 늦은 봄에 동일한 지역에서 A. anophagefferens의 대발생과 관계없이 P. ovalis의 높은 개체수는 보고되었다(Waterbury et al. 1986). P. ovalis의 대발생의 원인은 먹이원으로 추정되는 Synechococcus의 개체수 증가와 여과섭식 포식자의 부재에 의한 것으로 해석하였다(Johnson et al. 1988). Heterocapsa rotundata는 습도가 높고, 저수온기의 부영양화된 저염분수계에서 출현하는 것으로 보고되었다(Millette et al. 2015). 특히 우리나라는 서해안 시화호 및 새만금호에서 주로 겨울철 적조종으로 보고되고 있다(Seong et al. 2006). Millette et al. (2017)은 광영양부터 부분적 종속영양까지 환경에 따라 영양형의 전환이 가능한 H. rotundata의 혼합영양적 특성이 포식압이 낮고 다른 경쟁종의 생장이 낮은 환경에서 대발생을 야기할 수 있도록 한다고 제안하였다. 영랑호의 겨울철 낮은 수온, 부영양화, 낮은 포식압 등의 환경 조건이 H. rotundata의 적조 발생에 관련되었을 것이다.
수서생태계에서 식물플랑크톤의 개체수 및 밀도의 변화는 가용한 영양염, 경쟁 및 포식압 등의 다양한 생태학적 요인에 의해 결정된다(Reynolds and Reynolds 2006). 비록 본 연구에서 조사 당시 영양염 농도를 측정하지 못하였으나, 이전 영랑호 환경에 관한 보고서와 학술발표자료에 근거하여 2009년 영랑호에서 혼합 적조 발생은 영양염을 두고 경쟁하는 종들 사이에서 자원 경쟁을 넘어선 수준의 부영양화 상태가 진행되었을 것이라 추정된다. 또한 주요 우점한 3종의 영양형은 독립영양과 종속영양 두가지의 혼합영양을 보이는 종들로써 영양염 외에 부영양화의 영향을 받은 먹이원의 증가에 의한 것일 수도 있다. 즉, 겨울철 영랑호에서 혼합적조 발생은 우점종들 간의 연쇄적 상호 작용 보다는 각각의 종들이 서로 다른 생태적 지위를 갖기 때문에 가능했을 것으로 추정된다.
생태학에서 경쟁 배타원리에 반하는 플랑크톤 역설은 1961년 Hutchison에 의해 처음으로 제안되었고(Hutchinson 1961), 플랑크톤의 공존을 설명하기 위해 빛 또는 난류의 수직 기울기, 공생, 구별된 포식압과 끊임없이 변화하는 환경 등의 조건들이 제안되었다. 이후 플랑크톤 역설을 설명하기 위해 다양한 가설들[e.g. Chaotic mixing (Károlyi et al. 2000), 시공간적 이질성 (Miyazaki et al. 2006) 등 참조(Lee et al. 2015)]이 추가적으로 제안되고 있다. 비록 본 연구의 목적이 플랑크톤 역설 기작을 설명하기 위해 수행된 것이 아니기에 다양한 자료 분석이 수행되지 않아 영랑호의 혼합 적조 발생을 플랑크톤 역설에 준하여 해석하기에 섣부르나, 겨울철 석호의 혼합 적조 발생 현상의 향후 규명은 플랑크톤 역설을 설명하기 위한 추가적인 가능성을 제시해 줄 수 있을 것이다.
본 연구는 2009년 1월 동해안 대표 석호인 영랑호에서 겨울철에 발생한 식물플랑크톤 대발생에 관한 첫 기록이다. 특정한 1종의 우점에 의해 야기된 적조와 달리 단세포성 녹조류(Crpytomonas cf. marssonii), 와편모조류(Heterocapsa rotundata), 아메바류(Paulinella ovalis)의 3종에 의해 혼합 대발생이 발생된 것은 각 종의 생태적 지위의 차이에 의한 것이라 추정된다. 자연적, 인위적 변화를 수반하는 석호 생태계의 유지를 위해 지속적인 모니터링 조사는 기본적으로 수행되어야 하나, 이에 앞서 영랑호를 비롯한 동해안 석호에서 식물플랑크톤상에 관한 연구 자체는 전혀 수행된 바 없는 상황을 고려하여 담수와 해수의 중간 전이생태계에 대한 수생생물상의 연구는 우선적으로 선행되어야만 할 것이다.
