

1. 서론
홍조식물 곱슬이속(Plocamium)은 Lamouroux에 의해 1813년에 Plocamium vulgare J.V.Lamouroux를 기준종으로 하여 설립되었고(Lamouroux 1813), 전 세계 열대에서 추운 온대지역 해역까지 널리 분포한다. 곱슬이속은 전 세계에 약 45종이 보고되어 있으며(Guiry and Guiry 2022), 주로 조하대 암반이나 다른 엽상체 위에 착생하여 자라고 종종 부유하는 해조류(algae turf)에서 생육하기도 한다(Saunders and Lehmkuhl 2005). 곱슬이속은 연속 편생지(alternating series) 내 소지(branchlet)의 수, 연속 편생지 내 기저소지(lower branchlet)의 형태, 사분포자탁(tetrasporangial stichidia)과 낭과(cystocarp)의 형태 및 배열 등을 근거로 종을 구분하고 있다(Yano et al. 2004; Cremades et al. 2011).
Cremades et al. (2011)은 Dixon (1967)을 참고하여 Linnaeus가 관찰한 Van Royen의 표본(Herb. Lugdb. Bat.910.184.14)을 가는곱슬이(Plocamium cartilagineum)의 선정기준표본(lectotype)으로 지정하였으며 이를 북유럽(아일랜드, 프랑스) 및 스페인 표본과 함께 형태를 관찰하였다. 또한, Yano et al. (2004)은 연속 편생지 내 기저소지의 형태와 세포내 pH 농도를 각각 5가지, 3가지 유형으로 나누어서 종을 구분한 바 있다.
한국에 생육하고 있는 곱슬이속은 Kim et al. (2013)의 종목록에서 다음의 6종이 보고된 바 있다: 가는곱슬이(P. cartilagineum (Linnaeus) P.S.Dixon), 애기곱슬이(P. ovicorne Okamura), 뒷고리곱슬이(P. recurvatum Okamura), 수리곱슬이(P. suhrii Kützing), 참곱슬이(P. telfairiae (W.J.Hooker and Harvey) Harvey ex Kützing), 갈고리참곱슬이(P. uncinatum (C. Agardh) Kützing). 그러나 최근 곱슬이속에 대한 주요 식별 형질의 상사성(morphological homoplasy)이 알려지면서 이전의 종 분류에 대한 불완전성이 확인되었고(Yano et al. 2004, 2005) 아울러서 다수의 잠재종이 아직도 미기재 상태로 남아 있는 것으로 알려지고 있다(Kim et al. 2013; Saunders and Lehmkuhl 2005; Cremades et al. 2011). 즉 한국산 가는곱슬이(Plocamium cartilagineum)는 전 해역에 분포하지만 분류학적으로 연구가 미흡하고, 해조상 연구에서 출현 종 목록에 수록되는 정도에 그쳐 종의 실체 파악이 어렵다.
본 연구는 한국산 가는곱슬이의 엽상체 크기와 연속 편생지 내 소지의 수, 사분포자낭의 크기 등을 기준으로 형태를 분석함으로써 한국산 가는곱슬이의 분류학적 실체를 파악하고 원기재문과 비교·검토하여 가는곱슬이의 분류학적 한계를 설정하기 위하여 수행되었다. 더 나아가 한국산 가는곱슬이의 실체 파악 및 근연 분류군과의 유연관계를 명확히 하는 기초자료를 제공하는 것을 목적으로 한다.
2. 재료 및 방법
실험 재료는 한반도 동해, 남해, 제주도의 조하대 암반에서 착생하고 있는 엽상체 또는 떠밀려온 엽상체 등을 채집하였다(Table 1). 채집된 재료는 형태관찰을 위해 현장에서 3-5% 포르말린-해수로 고정하여 실험실로 운반하였다. 엽상체의 외부형태는 Olympus TG-4 방수카메라로 찍었다(Olympus, Tokyo, Japan). 내부구조 및 생식기관은 고정형 동결 마이크로톰(NK-101-II; Nippon Optical Works Co., Ltd., Tokyo, Japan)으로 박편을 만든 후, 1% aniline-blue 수용액으로 염색하고 해수로 씻은 후 35% 옥수수 시럽으로 고정하여 해부현미경(SZ51; Olympus)과 광학현미경(BX43; Olympus)으로 관찰하였다. 형태관찰을 위해 Yano et al. (2004)의 형질기준을 따라 엽상체의 윗부분(약 1 cm)을 대상으로 연속 편생지와 연속 편생지 내 기저소지를 Fig. 1과 같이 정의하였고 횡단한 주축의 폭(Fig. 1A)과 기저소지의 길이(Fig. 1B) 및 폭(Fig. 1C)을 광학현미경의 프로그램(CellSens Entry)을 이용하여 측정하였다.
Table 1.
Sample information of Plocamium cartilagineum collected from Korea.
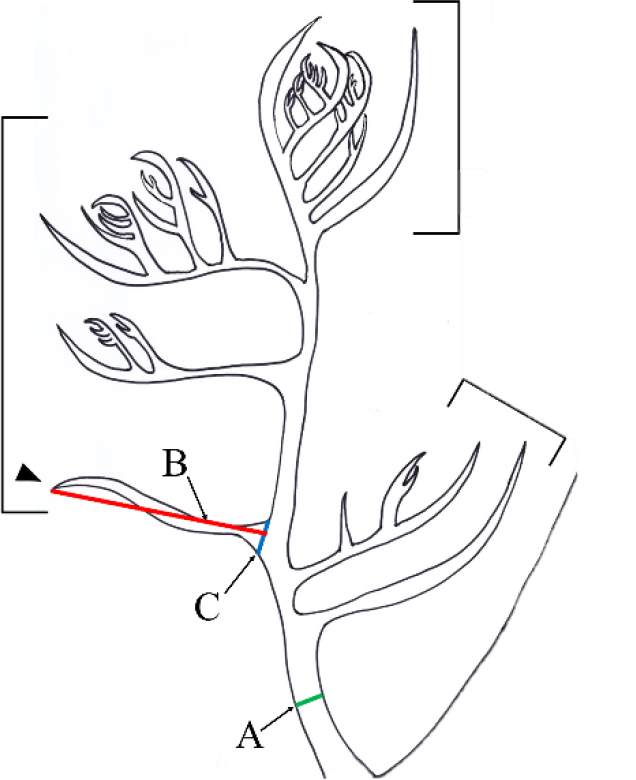
Fig. 1
Drawing apical branches of Plocamium cartilagineum showing the portion of morphological structures measured in this study. Arrowhead indicates the lowermost branchlets in each alternate series (square bracket). (A) the width of main axis. (B) the length of lower branchlet. (C) the width of lower branchlet. Scale bar: 2 mm.
관찰 결과는 EOS 600D 디지털 카메라(Canon, Tokyo, Japan)로 사진을 찍어 기록하였으며, 사진은 Adobe Photoshop 소프트웨어(ver. 6.1; Adobe Systems Inc., San Jose, CA, USA)를 이용하여 수정했고 건조표본은 제주대학교 생물학과 식물표본실(JNUB)에 보관하였다.
3. 결과
Plocamium cartilagineum (Linnaeus) P.S.Dixon 1967:
원기재명:Fucus cartilagineum Linnaeus
이명:Plocamium vulgare J.V.Lamouroux 1813; Plocamium cartilagineum var. uncinatum (C. Agardh) Guiry ex Benhissoune, Boudoureque & al. 2002; Plocamium uncinatum (C.Agardh) Kützing 1866; Fucus coccineus Hudson 1778
국명: 가는곱슬이
선정기준표본: 네덜란드 레이던(Herb. Lugdb. Bat. 910.184.14)
기준생육지: ‘In oceano australiore’ (more likely northern Europe fide Dixon 1967)
분포: 북대서양, 칠레, 북태평양 동부 및 서부, 아라비아해 북부, 호주, 뉴질랜드, 남극(Bischoff-Bäsmann and Wiencke 1996; Wynne 2002; Yano et al. 2004)
외부형태: 엽체는 진홍색 내지 밝은 홍색이고, 체장은 약 5.5 cm로 전체적으로 편압하며 섬유상 가근체로 부착하고 엽폭이 가는 띠 모양으로 연약하며 엉켜 자란다(Fig. 2A). 엽상체는 동일 평면상에 2-3개의 편생하는 소지가 한 단위가 되어 호생으로 배열하며 간혹 4개도 있다(Fig. 2B). 생장은 연기적으로 생장축이 바뀌면서 이뤄지고, 생장축은 하나의 세포로 이루어진 정단분열조직에서 유래한 단축구조로 주축은 식별 추적이 가능하다(Fig. 2B). 연속 편생지 내 기저소지는 분지하지 않고 단순하며 연변부가 매끈하고 기저부가 넓다. 또한 기저소지는 가늘고 선형이며 말단은 축 방향으로 굽고 끝이 날카로우며 상당히 길게 신장한다.
내부구조: 주축의 횡단면은 타원형이며 크기는 최대 2.3 mm에서 최소 0.3 mm로 평균 1.0 mm이다(Fig. 2C). 내부구조는 피층과 수층으로 나뉘며 수층은 중심세포와 주심세포로 구분되고(Fig. 2C), 하나의 중심세포는 4개의 주심세포를 형성하며(Kim and Hwang 2015) 이들 주심세포가 분화하여 위-유조직성 피층을 형성한다. 피층은 1-2개의 세포층으로 색소체를 함유한 작은 세포로 구성된다(Fig. 2C). 수층조직은 주심세포에서 피층세포까지 총 1-4개의 세포층으로, 거리가 짧은 곳은 주심세포가 1-2개, 긴 곳은 2-4개로 이뤄지며 수층조직은 크고 투명한 타원형 세포로 구성된다. 엽상체의 내부구조를 이루는 세포들의 크기는 다음과 같다. 피층세포의 폭은 최대 23.2 μm에서 최소 3.4 μm이며 평균 12 μm으로 긴 타원형이다(Fig. 2C). 단일 중심세포의 폭은 최대 75.3 μm에서 최소 45.4 μm로 평균 62 μm이며, 주심세포의 폭은 최대 121.5 μm이며 최소 46.7 μm로 평균 83 μm이고 중심세포와 같은 타원형이지만 크기가 더 크다.
생식구조: 자성배우체는 주축을 따라 낭과를 형성하며 성숙한 낭과는 구형이고 자루없이 형성되며 과공은 없다(Fig. 2D-2E). 과포자낭의 크기는 최대 43.2 μm에서 최소 18.1 μm로 평균 29.4 μm이다. 과포자는 조포사(gonimoblast) 끝부분에서 분열하여 다발을 형성하며 과포자끼리 방사형으로 연결되어 있다(Fig. 2F). 사분포자탁은 소지말단이 변형된 구조로(Fig. 2G), 초기에는 장상(掌狀)으로 분지하고(Fig. 2H) 성숙한 사분포자탁은 사분포자낭이 이열로 배열되며 이에 사분포자가 한 개씩 들어있다. 성숙한 사분포자는 대상(zonate)으로 분열하고 성장하면 사분포자탁이 신장되어 별-모양으로 발달한다(Fig. 2I). 영양체와 사분포자체는 여름~겨울까지 관찰되었으며 특히 과포자체는 7월과 11월 사이 관찰되었고 웅성배우체는 채집되지 않았다.
한국의 동해안과 남해안 연안에 분포하는 가는곱슬이는 ①엽상체의 크기와 ②주축의 폭(Fig. 1A), ③연속 편생지 내 기저소지의 크기(Fig. 1A-1C)를 측정하였으며 값은 다음과 같다. 동해안 엽상체의 크기는 최대 8 cm에서 최소 2 cm이고 평균 4 cm이며, 남해안 엽상체의 체장은 최대 25 cm에서 최소 7 cm이고 평균 10 cm이다. 동해안 엽상체의 주축의 폭은 최대 1.3 mm에서 최소 0.3 mm이고 평균 0.8 mm이며, 남해안 엽상체의 주축의 폭은 최대 0.6 mm에서 최소 0.04 mm이고 평균 0.4 mm이다. 제주에서 채집한 엽상체의 주축의 폭은 최대 0.4 mm에서 최소 0.2 mm이고 평균 0.3 mm이며 남해안과 값이 겹친다. 동해안 엽상체의 기저소지의 길이는 최대 1,806 mm에서 최소 242 mm이고 평균 844 mm이며, 남해안 엽상체의 기저소지의 길이는 최대 2,820 mm에서 최소 156 mm이며 평균 1,575 mm이다. 동해안 기저소지의 폭은 최대 729 mm에서 최소 130 mm이고 평균 370 mm이며, 남해안 엽상체의 기저소지의 폭은 최대 403 mm에서 최소 100 mm이며 평균 260 mm이다.
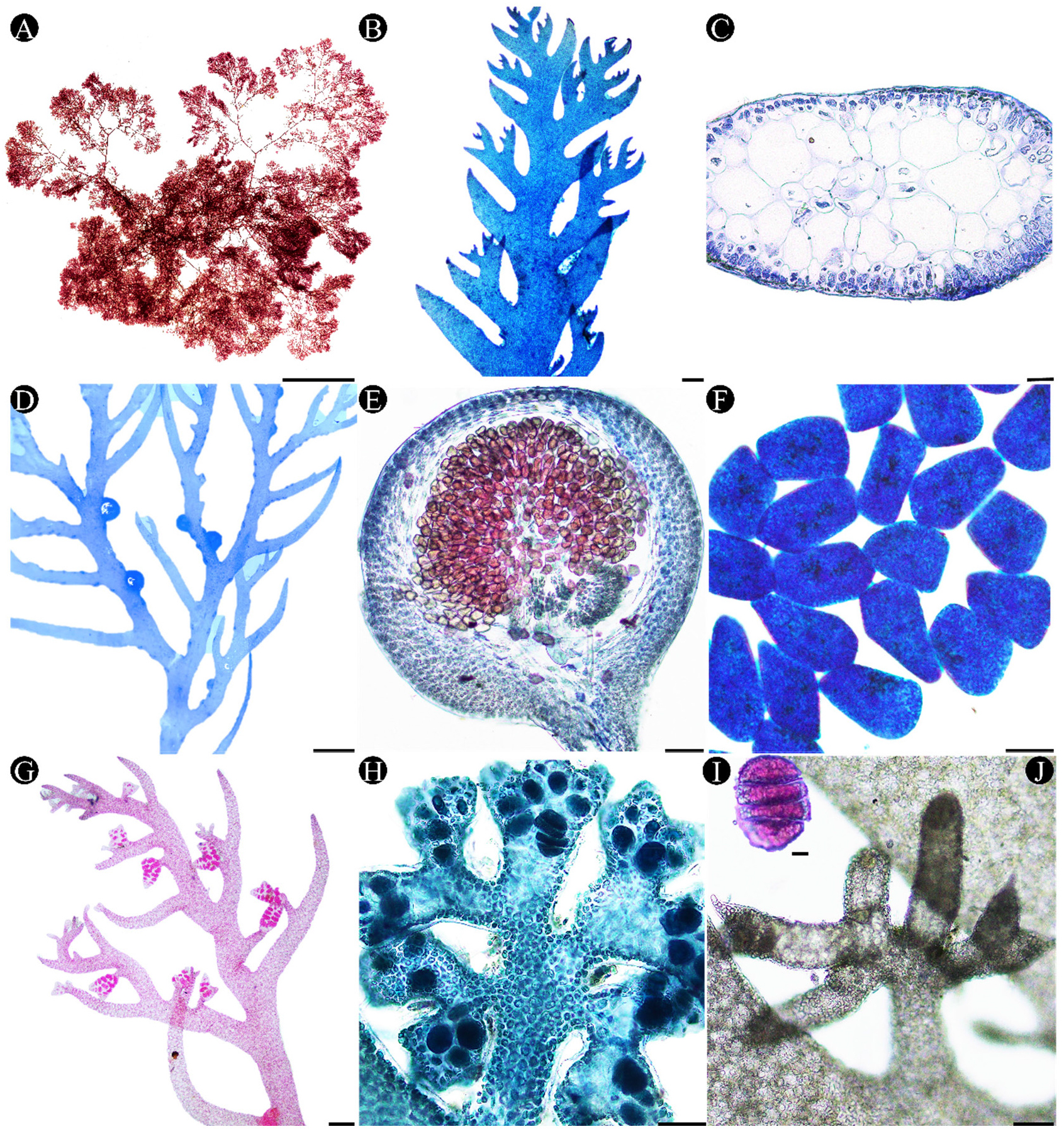
Fig. 2
Morphological structures of Plocamium cartilagineum. (A) Habit of thallus collected from Jongdal, Jeju, Korea on 3rd February 2013. (B) Apical part of branches showing alternate branchlets in series of 2~3 above the basal ramuli (C) Cross-section of a main axis showing subcylindrical shape. (D) Cystocarps along the main axis of a female gametophyte. (E) Cross section of cystocarp. (F) Mature carposporangium. (G) Stichidia on a tetrasporophyte. (H) Palma-shape of juvenile stichidia. (I) Tetrasporangia showing zonatae division. (J) Star-shape of old stichidia. Scale bars represent: A, 5 cm; B, 3 cm; C, 50 μm; D, 1 cm; E & J, 100 μm; F & H, 20 μm; G, 200 μm; I, 10 μm.
4. 고찰
곱슬이속에 속하는 식물종의 주요 식별형질은 ① 엽상체의 크기(height of thallus), ② 주축의 폭(width of main axis), ③ 연속 편생지 내 기저소지(lower branchlet)의 형태, ④ 연속 편생지 내 가지의 수(the number of branchlet per series), ⑤ 사분포자탁(tetrasporangial stichidia)의 형태, ⑥ 엽상체의 색, ⑦ 낭과의 형태 등으로 알려져 있다(Simons 1964; Womersley 1971; South and Adams 1979; Gabrielson and Scagel 1989). 본 연구에서는 가는곱슬이(P. cartilagineum)의 엽상체 색을 제외한 6가지의 형질을 측정하여 한국과 네덜란드, 일본, 유럽(스페인, 아일랜드, 프랑스) 개체군과 비교 분석하였다(Table 2).
Table 2.
Comparison of morphological characteristics of Plocamium cartilagineum from Korea, Japan, EUR2 (Spain, France, Ireland) and Netherlands as the type locality.
| Korea | Netherlands | EUR2 | Japan | |
| Height (cm) | 2.6 - 9.3 (5.5) | 11 | 5 - 20 (10) | - |
| Width of axes (mm) | 0.1 - 0.7 (0.6) | Up to 0.8 | 0.5 - 1.5 (1.0) | 0.4-1.5 |
|
Number of branchlet in a series | 2 or 3 (4) | 2 or 3 (4) | (2 or) 3 (or 4) | 3 or 4 (or 5) |
| Lower branchlet | ||||
| length (mm) | 0.55 - 2.24 (1.3) | Up to 1 | 0.5 - 4.5 (2.0) | 0.9 - 3.8 (1.5) |
| width (mm) | 0.05 - 0.55 (0.3) | Up to 0.25 | 0.10 - 0.50 (0.25) | 0.13 - 0.28 |
| ratio (L/W) | 1.4 - 11 (5.1) | Up to 8 | 4 - 14 (9) | Up to 7 |
| shape | Linear or adaxially curving | Slightly extrorse | Very falcate to extrorse |
Most of linear, few adaxially curving or slightly recurved |
| Cystocarp size | 0.3 mm in diameter | 1 mm in diameter | Up to 1 mm in diameter | - |
| Tetrasprangium size | Up to 67 μm in length | Up to 80 μm in length | 70 - 90 (80) μm in length | - |
| Stichidia | ||||
| shape(juvenile) |
Terminally lot of furcated or not, narrow stalk |
Simple or terminally bifurcated, narrow stalk |
Simple or terminally bifurcated, rarely trifurcated, with a neat stalk | - |
| shape(mature) |
Complexed stichidia have a long stalk, bifurcated stichidia dichotomously branched from the middle or tip of their length |
Simple stichidia have a long stalk, bifurcated stichidia dichotomously branched from the middle of their length |
Simple stichida have a long stalk, bifurcated stichidia dichotomously branched from the middle of their length | - |
| Reference | This study | Cremades et al. (2011) | Cremades et al. (2011) | Yano et al. (2004) |
Cremades et al. (2011)은 주요한 식별형질(①-⑦)을 바탕으로 가는곱슬이(Plocamium cartilagineum)의 선정기준표본(lectotype)과 스페인, 아일랜드, 프랑스산을 포함하는 EUR2 표본을 관찰하였다. 가는곱슬이의 선정기준표본과 EUR2을 종합한 결과, 가는곱슬이의 특징으로 엽체는 암적색을 띠고 연속 편생지 내 소지는 2개에서 3개이며 말단으로 갈수록 좁아지고 끝이 약간 밖으로 향하며 갈고리처럼 굽어 있음을 기재하였다. COI-5P 미토콘드리아 유전자를 대상으로 한 분자계통학적 연구에서 아일랜드, 프랑스, 스페인 갈리시아(Galicia)산은 같은 하플로타입(haplotype)을 가지고, 스페인 카탈루냐(Catalonia)와 안달루시아주(Andalusia)는 3개의 하플로타입을 나타내었으며 세 나라 내 염기서열 변이 차이는 0.2-0.5%으로 보고하였다(Cremades et al. 2011).
본 연구에서 관찰한 한국산 가는곱슬이는 Cremades et al. (2011)가 관찰한 형질에 따라 ① 엽상체 크기, ③ 연속 편생지 내 기저소지의 형태, ⑤ 사분포자탁(stichidia)의 형태, ⑦ 낭과의 형태에서 기준 생육지인 네덜란드산과 차이가 난다. 한국산은 네덜란드산에 비해 엽상체의 크기가 2배 가량 작으며 한국산의 기저소지는 네덜란드산보다 길고 폭이 넓으며 길이와 폭의 비가 작다. 한국산의 연속 편생지 내 기저소지는 축방향으로 끝이 살짝 굽은 선형이며, 네덜란드산은 끝이 밖으로 구부러진다(Table 2). 네덜란드산의 사분포자탁은 끝이 이분지되거나 분지되지 않지만, 한국산의 어린 사분포자탁은 이분지되거나 장상(掌狀)으로 분지되고(Fig. 2H) 성숙한 사분포자탁은 별-모양으로 분지한다(Fig. 2J). 낭과의 크기는 한국산이 네덜란드산에 비해서 약 3배 작으며 연속 편생지 내 소가지의 수와 주축의 폭은 비슷하다(Cremades et al. 2011).
Yano et al. (2004)은 일본산 곱슬이속(P. telfairiae, P. recurvatum, P. cartilagineum, P. ovicorne, P. serratulum)을 대상으로 형태적인 차이와 생리적인 특징을 기초로 하여 구분하였다. 연속 편생지 내 기저소지의 크기 및 기저소지의 굽은 형태에 따라 5가지 유형(adaxially curving, linear, slightly recurving, strongly recurving, coiling)으로 나누고 세포 내 pH 농도로 종의 특징을 설명하였다. 일본산 가는곱슬이는 3-4개의 연속 편생지를 가지며 기저 소지 형태는 대개 선형이고 종종 축을 향해 굽은 형(adaxially curving)이다. 또한 엽록체 루비스코(rubiso) 염기서열을 분석한 결과 3개의 하플로타입(haplotypes)을 확인하였으며 그 중 1개는 강산(strong acidity)이며 나머지 두 개는 중성의 pH를 가진다고 보고하였다.
본 연구에서 관찰한 한국산 가는곱슬이는 Yano et al. (2004)이 관찰한 형질에 따라 ① 주축의 폭, ② 연속 편생지 내 기저소지의 형태 및 길이와 폭, ③ 연속 편생지 내 소지의 개수에서 일본산과 차이가 난다(Table 2). 한국산은 일본산보다 주축의 폭이 좁고 연속 편생지 내 기저소지는 약간 짧으며 넓다. 특히 기저소지의 폭의 경우 한국산은 최대값이 0.5 mm인 반면, 일본산은 최대값이 0.28 mm로 한국산보다 다소 좁다. 연속 편생지 내 소가지의 수는 일본산이 3-4개로 한국산보다 분지가 많이 되며, Yano et al. (2004)의 분류에 따라 연속 편생지의 형태는 한국산과 일본산이 비슷하나 일본산은 밖으로 굽는 형이 관찰된다.
한국의 동해안과 남해안 연안에서 채집한 엽상체를 관찰한 결과, ① 엽상체의 크기, ② 주축의 폭, ③ 기저소지의 크기에 차이가 있음을 확인하였는데 이것은 생육환경에 따른 형태변이로 추정된다(Cremades et al. 2011). 파고가 높은 동해안은 남해안에서 채집한 엽상체들에 비해 엽상체의 크기가 작고(Fig. 3A-3B) 주축의 폭이 2배가량 넓으며(Fig. 3C), 파고가 낮은 남해안은 동해안에서 채집한 엽상체들과 비교했을 때, 연속 편생지 내 기저소지의 폭이 좁고 길이가 약 2배 길다(Fig. 3D). 유럽산(EUR2)을 한국산과 비교했을 때, 유럽산은 주축의 폭이 동해안과 유사하며 한국산과 0.6 mm이하에서 형질의 불연속성을 나타내고, 기저소지의 크기는 남해안과 유사하며 유럽산은 최대 4.5 mm로(Table 2) 한국산보다 길고 0.3 mm이상에서 형질의 불연속성을 보이므로 분자계통학적인 방법론을 도입하여 한국산과 기준생육지인 네덜란드산에 대한 종의 실체를 명확하게 확인해야 한다.
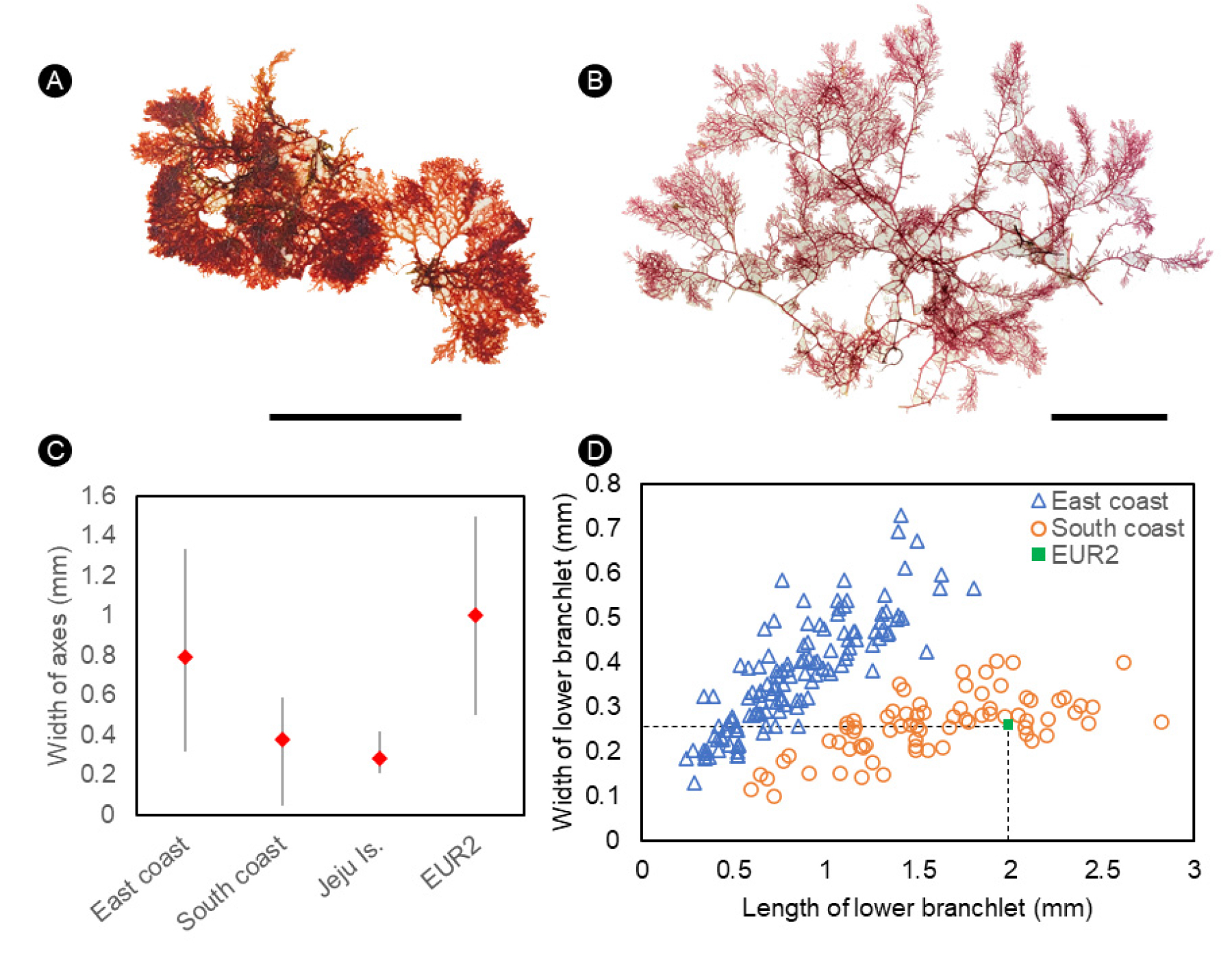
Fig. 3
Morphological comparison of Plocamium cartilagineum collected from the East and South coasts of Korea. (A) Habit of thallus from Donghea, Cheongok-dong, Korea on 24th Octorber 2020. (B) Habit of thallus from Yeosu, Deokchon-ri, Korea on 12th October 2016. (C) Variation of the width of main axis in the cross section of specimens from the East and South coasts of Korea, Jeju Island, and specimens of EUR2 (Spain, France, Ireland) from Europe (Creamades et al. 2011). (D) Variation of the length and width of the lower branchlet in the specimens from the East and South coasts of Korea, and the average of EUR2 (Spain, France, Ireland) from Europe (Creamades et al. 2011). Scale bars: A & B, 3 cm.
본 연구 결과 한국산 가는곱슬이는 기부에서부터 분지가 시작되며 연속 편생지 내 소지의 수는 2개에서 3개이고 편생지 내 기저소지는 말단으로 갈수록 가늘어지며 개체군내에서 형태변이를 보인다. 동해안표본은 남해안표본에 비해 전체적으로 엽상체의 크기가 작고 주축의 폭이 넓으며 기저소지가 짧고 폭이 넓다. 낭과는 주축 가장자리를 따라 자라며 네덜란드산과 달리 낭과의 크기가 작고 사분포자탁이 장상(掌狀)으로 자라다가 별-모양으로 발달한다. 일본산과 비교하면 한국산은 기저소지의 길이가 짧고 폭이 매우 넓다.
최근 가는곱슬이에서 추출한 할로겐화된 모노테르펜류(monoterpenes)는 사람의 암세포(CT26, SW480, HeLa)에 대하여 세포독성 효과가 있는 것으로 밝혀졌다(de Inés et al. 2004). 이러한 생물자원으로의 활용 가능성이 있는 가는곱슬이는 지리적 분포와 생육환경에 따른 종 내 형태적 차이를 보이므로 종을 정확하게 동정하기위해 DNA 염기서열에 근거한 보다 명확한 종 동정 기준이 필요하다. 앞으로 추가연구에서 기준지인 네덜란드와 일본, 한국 지역집단간의 분자계통학적 유연관계를 확인하여 이들의 계통분류학적 위치를 재검토하여야 할 것이다.
