Copyright © 2026 The Korean Society of Phycology
ABSTRACT
1. 서론
2. 김 유리사상체의 확보
3. 김 유리사상체 배양
4. 김 유리사상체 연구 동향
5. 국내 김 품종개발 현황
6. 결론
1. 서론
홍조류 김(Pyropia spp.)은 한국, 일본, 중국 등 동아시아 지역에서 오랜 기간 양식되어 온 해조류이다(He et al. 2021; Kim et al. 2022). 국내 연안에서는 20여종이 서식하고 있으며, 상업적으로 양식하고 있는 김은 방사무늬김(Pyropia yezoensis), 잇바디돌김(Pyropia dentata), 모무늬돌김(Pyropia seriata) 3종으로, 일본(방사무늬김)과 중국(방사무늬김, 하이타넨시스김) 등 주요 김 생산국과 비교하면 상대적으로 다양한 김 종이 양식 산업에 활용되고 있다(Sasuga et al. 2017; Ding et al. 2021). 과거에는 참김(Pyropia tenera) 또한 양식되었다고 알려져 있으나 현재는 거의 사라지고(Hwang et al. 2005, 2010), 환경적응력과 생산성이 탁월한 방사무늬김의 생산량이 국내 김 생산량의 70% 이상을 차지하고 있다.
김의 생활사는 육안으로 관찰되는 엽상형 배우체(gametophytic phase, n)와 현미경적 크기인 사상형 포자체(sporophytic phase, 2n)가 이형세대교번(heteromorphic alternation of generations)을 한다(Luo et al. 2014; Kim et al. 2022). 사상체(conchocelis)는 엽상체와 형태적으로 현저한 차이를 보여 1949년 이전까지는 Conchocelis rosea라는 독립된 종으로 기록되었다. 이후 Drew (1949)에 의해 사상체가 김의 생활사 단계임이 밝혀졌으며, 사상체는 각포자(conchospore)를 형성하고 방출된 각포자는 발아하여 엽상체로 성장한다(Fig. 1).
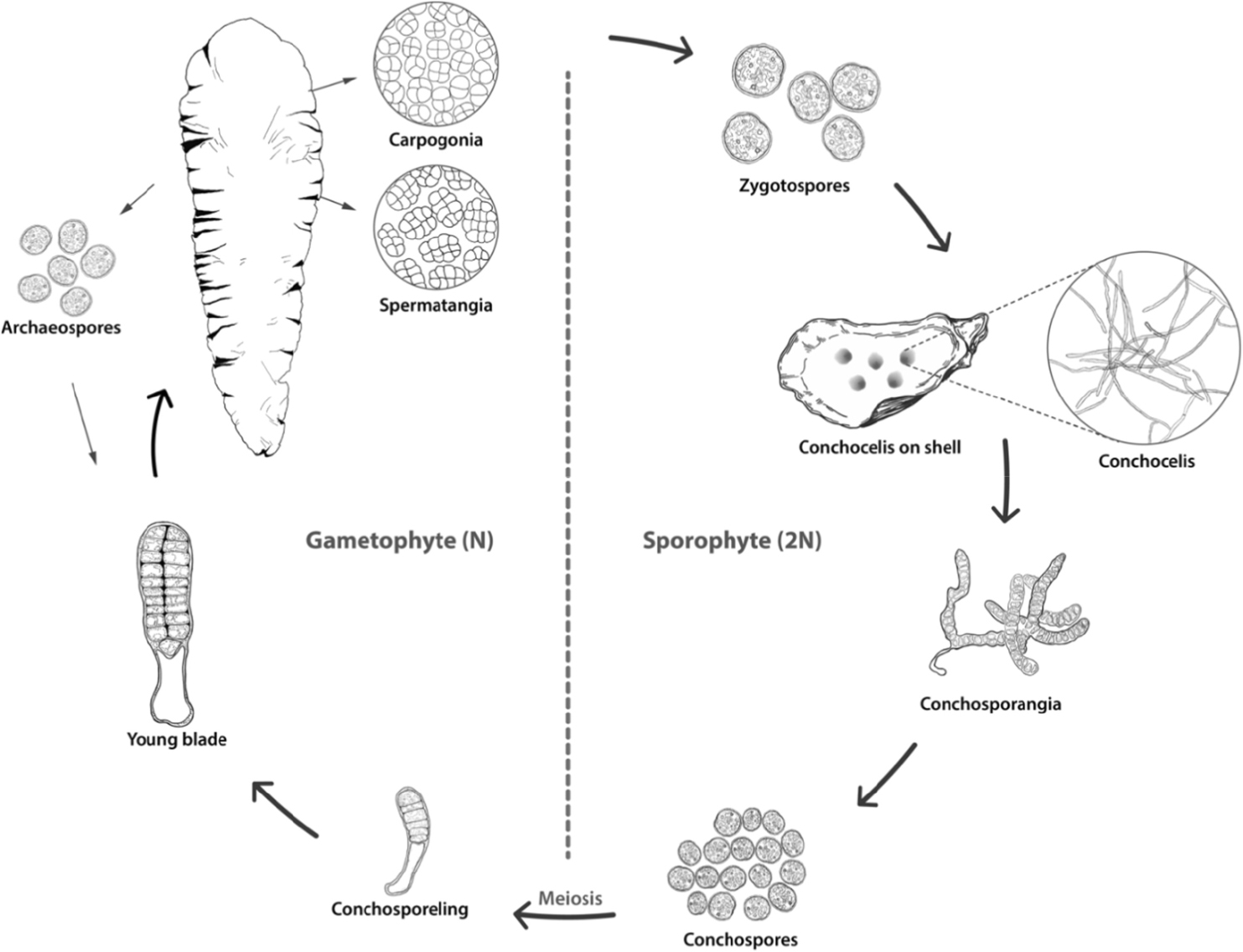
Fig. 1
Pyropia heteromorphic life cycle alternates between gametophytic stage and filamentous sporophyte stage to Kim et al. (2022).
김 사상체는 일반적으로 굴 껍데기와 같은 패각 내부에 잠입하여 생장하는 패각사상체(shell-living conchocelis)의 형태로 존재하지만, 실내 배양 환경에서는 기질에 부착하지 않고 자유롭게 생장하는 유리사상체(free-living conchocelis) 형태로도 유지될 수 있다(He and Yarish 2006). 유리사상체는 계대배양을 통해 장기간 보존과 대량 증식이 가능하며, 증식된 유리사상체는 패각에 이식하여 김 양식망의 채묘에 사용되는 패각사상체 생산에 이용된다. 이러한 기술은 우량한 형질을 가진 계통주의 지속적인 유지와 안정적인 종자 생산을 가능하게 하여 김 양식 산업 발전의 기반이 되어 왔다(Blouin et al. 2011). 최근 기후변화에 따른 수온 상승과 양식환경 변화로 인해 김 양식 산업의 불안정성이 증가하고 있으며, 이에 따라 안정적인 종자 생산과 고수온 등 환경 변화에 대응 가능한 품종 개발의 중요성이 높아지고 있다. 유리사상체는 유전자원 보존, 생리·생태 연구 및 품종 개발을 위한 육종 소재로 활용되는 등 다양한 분야에서 중요한 역할을 수행하고 있으며(Jiang et al. 2020; He et al. 2021; Yan et al. 2024), 최근에는 기후변화 대응 품종 개발과 안정적인 종자생산 체계 구축을 위한 핵심 소재로 활용되고 있다(Lee et al. 2019; Heo et al. 2024). 따라서 본 논문에서는 유리사상체의 확보 방법, 배양 기술 및 연구 동향을 간략히 소개하여 김 유리사상체의 효율적인 활용과 종자 산업 발전을 위한 기초자료를 제공하고자 한다.
2. 김 유리사상체의 확보
유리사상체(free-living conchocelis)는 패각 등 탄산칼슘 기질에 부착하지 않고 배양액 내에서 자유롭게 생장하는 사상체를 말하며(He and Yarish 2006), 실내 배양 환경에서 인위적으로 관리 및 증식시킬 수 있다. 유리사상체는 가는 사상형 세포가 반복적으로 분지하는 형태를 가지며, 배양 초기에는 단순한 사상 구조를 나타내지만 생장이 진행됨에 따라 복잡한 망상 구조로 발달한다(Fig. 2E-G) (Hiwatashi et al. 2022). 또한 세포 내 엽록체를 이용한 광합성을 통해 생장하며, 분지를 반복하면서 방사형 둥근 형태의 군체(colony)를 형성한다(Fig. 2H-I) (Iwasaki 1961; Lee and Fultz 1970).

Fig. 2
Isolation and culture process of free-living conchocelis from Pyropia thalli; preparation of specimens (A), excision and washing of a portion of mature thalli (B-C), released carpospores (D), formation of conchocelis from carpospores (E), free-living conchocelis filaments (F-G), and culture of free-living conchocelis (H-I).
유리사상체는 현장에서 채집한 성숙한 김 엽상체로부터 방출된 과포자(carpospore)를 이용하여 확보한다(Fig. 2). 과포자는 성숙한 암배우체의 과포자낭(carposporangium)에서 형성되는 이배체 포자로, 방출 후 발아하여 사상체로 생장한다(Drew 1949; Cole and Conway 1980). 김의 유리사상체를 확보하기 위해 자연 서식지 또는 양식장에서 형질이 우수하거나 특성이 다른 개체를 선별한 후 성숙 여부를 확인한다(Fig. 2A). 미성숙 엽체의 경우 수온 20℃, 광량 40 μmol photons m–2 s–1 및 광주기 14L:10D 조건에서 배양하여 성숙을 유도할 수 있다. 성숙한 엽체는 과포자낭이 형성된 부위를 절단(1×1 cm)하여 페트리디쉬 또는 배양 용기에 넣고 멸균해수 또는 여과해수로 세척하여 부착된 퇴적물과 오염생물을 제거한 후 과포자 방출을 유도한다(Fig. 2B-C). 방출된 과포자는 발아 후 세포 분열과 분지를 반복하면서 유리사상체로 생장하고(Fig. 2D-G), 약 3~4주 후에 오염 여부를 확인하여 건강한 사상체를 선별한 후 PES 멸균해수(Provasoli 1968)에서 배양한다(Fig. 2H-I). 과포자를 이용한 방법은 유전적으로 안정적인 배양주 확보에 유리하며, 현재 김 유리사상체 확보에 가장 널리 이용되는 방법으로 알려져 있다(He and Yarish 2006). 확보된 유리사상체는 종자 생산을 위한 원종 유지, 유전자원 보존 및 품종 개발을 위한 육종 연구의 기초 소재로 활용된다.
3. 김 유리사상체 배양
유리사상체의 생장과 성숙은 온도, 광량, 광주기 및 영양염과 같은 다양한 환경 요인의 영향을 받으며(He and Yarish 2006; Li et al. 2011; Lu and Yarish 2011; He et al. 2021), 실내 배양 환경에서는 이러한 조건을 인위적으로 조절함으로써 생장과 성숙 단계를 제어할 수 있다. 일반적으로 영양생장 단계에서는 지속적인 세포 분열과 분지에 의해 군체 크기가 증가하며, 환경 조건의 변화에 따라 각포자낭(conchosporangia) 형성과 같은 생식 생장 단계로 전환된다. 이러한 생장 및 성숙 반응은 종에 따라 차이를 보이며, 각 종별 최적 배양 조건이 보고되어 있다(Table 1). 방사무늬김(Pyropia yezoensis)의 경우 유리사상체 최적 생장 조건은 수온 15~20℃로 보고되었으며(Seo et al. 2025), 각포자낭 형성 최적 조건은 수온 25℃, 광량 57 μmol photons m–2 s–1, 광주기 8L:16D로 보고되었다(Li et al. 2011). 잇바디돌김(P. dentata) 유리사상체는 수온 20~25℃와 광량 1~80 μmol photons m–2 s–1 범위에서 안정적인 생장을 보이며(Kim 1999), 특히 수온 25℃ 이하의 장일 조건(≥14L)에서 영양생장이 촉진되는 것으로 알려져 있다(Zhong et al. 2016). 반면, 각포자낭 형성은 수온 28℃, 광량 40~60 μmol photons m–2 s–1 및 광주기 12L:12D 조건에서 가장 높은 수준을 나타내었다(Zhong et al. 2016). 모무늬돌김(P. seriata) 유리사상체의 최적 생장 조건은 15~20℃, 20~80 μmol photons m–2 s–1로 보고되었으며, 각포자낭 형성은 20~25℃에서 활발하게 나타났다(Kim and Notoya 2004). 이와 같이 유리사상체의 생장 및 성숙 조건은 종에 따라 차이를 보이므로, 배양 목적과 대상 종에 따라 적절한 환경 조건을 설정하는 것이 중요하다. 예를 들어, 방사무늬김과 모무늬돌김은 비교적 낮은 수온(15~20℃)에서 생장이 우수한 반면, 잇바디돌김은 상대적으로 높은 수온(20~25℃)에서도 안정적인 생장을 나타내어 종별 생리적 특성의 차이를 보여준다. 또한 영양 생장에 적합한 조건과 각포자낭 형성에 적합한 조건이 서로 다른 경우가 많아, 유리사상체의 증식과 성숙 유도를 위해서는 배양 단계별 환경조건 조절이 필요하다. 이러한 배양 기술은 유리사상체의 대량 증식과 안정적인 원종 관리에 활용될 수 있으며, 종자 생산 효율 향상에 중요한 역할을 한다.
Table 1
Optimal culture conditions for vegetative growth and conchosporangia formation of free-living conchocelis in Pyropia species.
|
Species
|
Optimum conditions
|
References
|
|
Growth
|
Conchosporangia formation
|
| Pyropia yezoensis |
15~20°C, 100 μmol photons m–2 s–1 |
20~25°C, 57~100 μmol photons m–2 s–1,
8L:16D
| Li et al. (2011);
Seo et al. (2025) |
| Pyropia dentata |
20~25°C, 40 μmol photons m–2 s–1,
≥ 14L
|
25~28°C, 20~60 μmol photons m–2 s–1,
≥ 12L
| Kim (1999);
Zhong et al. (2016) |
| Pyropia seriata |
15~20°C, 20~80 μmol photons m–2 s–1,
|
20~25°C
| Kim and Notoya (2004) |
| Pyropia pseudolinearis |
20°C, 40 μmol photons m–2 s–1 |
10~25°C, 10~80 μmol photons m–2 s–1 | Kim (1999) |
| Neopyropia katadae |
16~20°C, 60~80 μmol photons m–2 s–1 |
–
| Liang et al. (2022) |
| Porphyra leucosticta |
15°C, 40 μmol m–2 s–1, 16L:8D
|
20°C, 40 μmol m–2 s–1, 8L:16D
| Ott (1965);
He and Yarish (2006) |
유리사상체 배양에는 일반적으로 Provasoli’s Enriched Seawater (PES) 배지(Provasoli 1968) 또는 Modified Grund Medium (MGM) 배지(McLachlan 1973)가 사용된다. 두 배지 모두 질산염, 인산염 등 무기 영양염과 미량 금속, 비타민류가 첨가된 멸균 해수 기반의 배지로, 배지는 영양염 부족 및 노폐물 축적을 방지하기 위해 배양액을 주기적으로 교환한다.
유리사상체의 안정적인 유지와 증식을 위해서는 계대배양과 오염 관리가 필요하다. 배양 과정에서 발생하는 사상 남세균(cyanobacteria) 및 기타 미생물의 오염은 유리사상체의 생장을 저해하는 주요 요인으로 알려져 있으며(Deng et al. 2022; Ma et al. 2023), 오염이 발생하면 건강한 사상체를 선별하여 계대배양하거나 필요에 따라 streptomycin, penicillin 등의 항생제 처리가 활용되기도 한다(Liu et al. 2002). 계대배양은 일반적으로 3~4주 간격으로 수행하며, 크기가 커진 군체를 핀셋이나 균질기를 이용하여 사상체를 세절한 후 새로운 배지에 분주하여 배양한다. 또한 장기간 계대배양 시 사상체의 활력이 저하되거나 형태적 변화가 나타날 수 있으므로, 사상체의 상태를 정기적으로 점검하고 건강한 사상체를 유지하기 위한 관리가 필요하다.
4. 김 유리사상체 연구 동향
김 유리사상체에 관한 연구는 생활사 규명과 배양 기술 확립을 시작으로(Drew 1949; He and Yarish 2006; Blouin et al. 2011), 유전자원 보존, 육종 및 분자생물학 연구 등으로 점차 다양화되고 있다(Jiang et al. 2020; Lin et al. 2021; Deng et al. 2022). 다양한 종 및 계통의 유리사상체를 장기간 보존하여 연구 및 종자 생산을 위한 원종으로 활용하고 있으며, 오염 관리 및 정제 기술 연구도 함께 이루어지고 있다(Deng et al. 2022; Ma et al. 2023).
또한 온도, 광량 및 영양염 변화에 따른 생장과 성숙 반응, 광합성 특성 및 환경 스트레스 반응을 규명하기 위한 연구가 지속적으로 수행되고 있으며(Li et al. 2011; Kim et al. 2019; He et al. 2021; Seo et al. 2025), 유리사상체와 엽상체가 고온 스트레스에 서로 다른 생리적 반응을 나타내는 것으로 보고되어, 생활사 단계별 환경 적응 기작과 이형세대교번의 생태학적 의미를 규명하기 위한 연구에도 활용되고 있다(Luo et al. 2014). 최근에는 전사체 및 유전체 분석을 통한 유전자 기능 분석 연구가 증가하고 있으며(Wang et al. 2020; Lin et al. 2021; Huang et al. 2025), 유리사상체 자체의 기능성 소재 및 바이오 소재로서 활용 가능성도 제시되고 있다. 이처럼 유리사상체를 활용한 연구는 단순한 생장 특성 규명에서 환경 스트레스 적응 기작의 해석과 유전자 수준의 기능 분석으로 확대되고 있다. 유리사상체는 실내에서 장기간 유지가 가능하고 환경 조건을 정밀하게 제어할 수 있어 이러한 연구를 수행하기 위한 유용한 실험 소재로 활용되고 있다.
이와 함께 유리사상체를 이용한 패각사상체 생산 및 종자 생산 기술은 안정적인 종자 공급을 위한 핵심 기술로 활용되고 있으며(He and Yarish 2006), 최근에는 패각 잠입 조건과 종자 생산 효율 향상을 위한 연구도 지속적으로 수행되고 있다(Heo et al. 2021, 2024). 또한, 유리사상체를 활용하여 선발육종과 돌연변이 육종을 통한 우량계통주 선발 및 품종 개발 연구가 수행되고 있으며, 최근에는 고수온 내성 품종 개발을 위한 연구가 활발히 진행되고 있다(Shin et al. 2018; Lee et al. 2019; Park 2023; Yan et al. 2024).
5. 국내 김 품종개발 현황
국내 김 품종개발은 선발육종, 교잡육종 및 돌연변이(감마선) 육종을 중심으로 이루어지고 있으며(Kim 2001; Park and Hwang 2014; Lee et al. 2019), 2012년 수산식물 품종보호제도 도입 이후 김 품종 개발과 품종 보호가 본격적으로 추진되었다(Park et al. 2016; Hwang and Park 2020). 현재까지 품종보호권이 등록된 김 품종은 국가기관(국립수산과학원) 17품종, 지자체 9품종, 산업계 2품종, 대학 1품종 및 정부출연연구기관 1품종으로 총 30품종이다(Fig. 3). 이처럼 국내 김 품종 개발은 공공 연구기관을 중심으로 추진되어 왔으며, 생산성 향상을 위한 속성장 품종 개발이 주를 이루었다(Hwang and Park 2020). 김 품종보호권의 등록 및 출원 현황은 국립수산과학원 e-연구바다(https://www.nifs.go.kr/portal)의 수산식물품종보호 메뉴에서 확인할 수 있다.
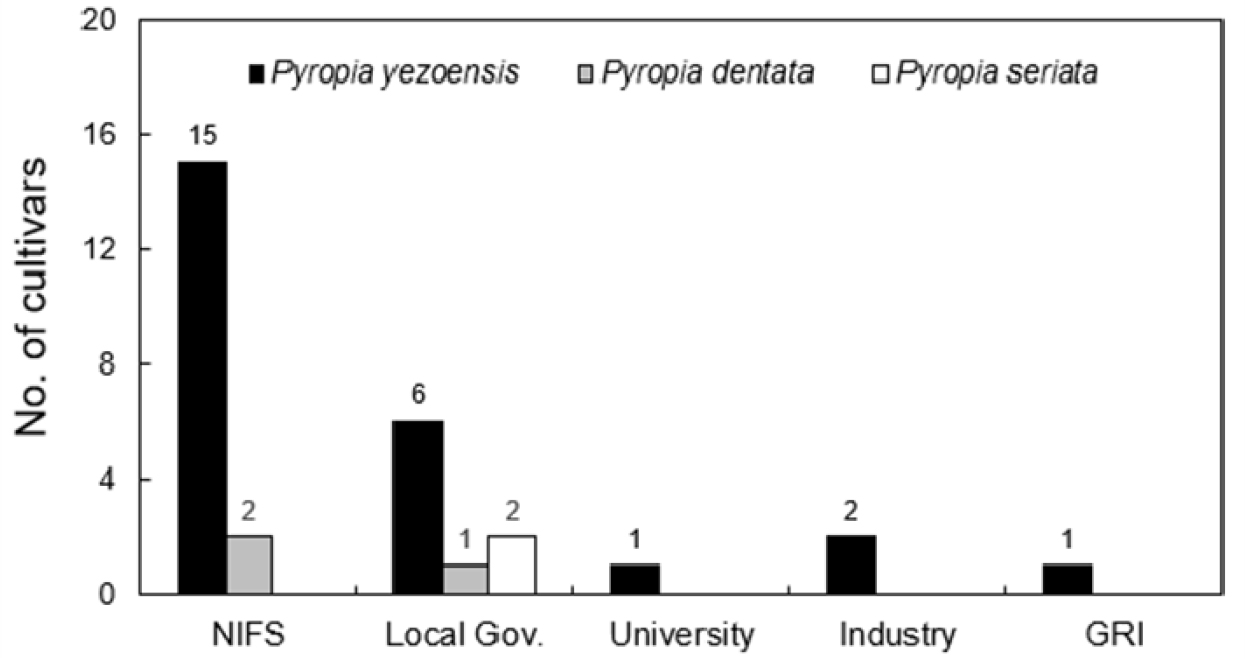
Fig. 3
Registration status of Pyropia cultivars in Korea. NIFS, National Institute of Fisheries Science; GRI, Government-funded Research Institute.
이러한 품종 개발의 지속적인 추진을 위해서는 다양한 유전자원의 확보와 체계적인 관리가 필수적이다. 유리사상체는 계대배양을 통해 장기간 보존이 가능하고 우량한 형질 특성을 안정적으로 유지·관리할 수 있으므로 품종 개발을 위한 유전자원 관리 측면에서중요한 역할을 한다. 이에 국립수산과학원은 다양한 종 및 계통의 김 유리사상체를 보존·관리하고 있으며, 품종개발, 종자생산 등을 위한 자원으로 활용하고 있다. 현재까지 등록·보존 중인 김 유리사상체는 11종 188계통주로, 방사무늬김 114계통주, 잇바디돌김 42계통주, 모무늬돌김 18계통주가 포함되어 있으며, 보존 중인 계통주는 연구 활성화를 위해 필요 시 외부 기관에 분양되고 있다. 보존 중인 유리사상체의 정보는 국립수산과학원 e-연구바다(https://www.nifs.go.kr/portal)의 수산생명자원정보센터에서 확인할 수 있다.
6. 결론
유리사상체는 김 생활사에서 사상형 포자체 단계에 해당하며, 실내 배양 환경에서 장기간 유지와 대량 증식이 가능하다. 유리사상체는 주로 과포자를 이용하여 확보하며, 온도, 광량 및 광주기 등의 환경 조건을 조절함으로써 안정적인 배양과 성숙 유도가 가능하다. 또한 유전자원 보존, 육종 연구 등 다양한 분야에서 활용되고 있으며, 김 양식 산업의 안정적인 종자 생산뿐만 아니라 기후변화 대응 품종 개발을 위한 핵심 소재로 이용되고 있다. 본 논문에서는 유리사상체의 확보 방법과 배양 기술, 그리고 주요 활용 분야를 간략히 소개하였으며, 관련 연구와 종자 생산 현장에서 유리사상체에 대한 이해를 높이기 위한 기초자료로 활용되어 안정적인 종자 생산 기반 구축과 김 산업의 지속가능한 발전에 기여할 것으로 기대한다.
Acknowledgements
본 연구는 국립수산과학원 해조류연구소 「기후변화 대응 김 종자생산 고도화 및 고수온 내성 품종 산업화 연구, R2026021」 과제의 일환으로 수행되었습니다.
References
Deng Y, Tian C, Hu C, Xu G, Yang L, Lu Q and Zhou W. 2022. The identification of filamentous cyanobacteria isolated from Neopyropia germplasm bank illustrates the pattern of contamination. J. Mar. Sci. Eng. 10, 838. https://doi.org/10.3390/jmse10060838.
10.3390/jmse10060838He B, Niu J, Xie X and Wang G. 2021. Development of free-living sporangial filaments regulated by light and culture density in Neopyropia yezoensis. Algal Res. 58, 102378. https://doi.org/10.1016/j.algal.2021.102378.
10.1016/j.algal.2021.102378He P and Yarish C. 2006. The developmental regulation of mass cultures of free-living conchocelis for commercial net seeding of Porphyra leucosticta from Northeast America. Aquaculture 257, 373-381. https://doi.org/10.1016/j.aquaculture.2006.03.017.
10.1016/j.aquaculture.2006.03.017Heo JS, Kim JK and Choi HG. 2024. Effects of environmental and physical factors on the shell infiltration density of free-living conchocelis of three Pyropia species (Bangiales, Rhodophyta). Algae 39, 319-327. https://doi.org/10.4490/algae.2024.39.12.14.
10.4490/algae.2024.39.12.14Heo JS, Park EJ, Hwang MS and Choi HG. 2021. Effect of shell-type, light and temperature on the shell infiltration of free-living conchocelis of three Pyropia species. Korean J. Fish. Aquat. Sci. 54, 23-30. https://doi.org/10.5657/KFAS.2021.0023.
10.5657/KFAS.2021.0023Hiwatashi Y, Shimada M, Mikami K and Takada N. 2022. Establishment of a live-imaging analysis for polarized growth of conchocelis in the multicellular red alga Neopyropia yezoensis. Front. Plant Sci. 12, 716011. https://doi.org/10.3389/fpls.2021.716011.
10.3389/fpls.2021.71601135251057PMC8888420Huang X, Wang Y, Li F, Zhao H, Zeng L, Li H, Gu F, Tan D, Hu W, Guo A, Ji C and He L. 2025. Physiological activities, transcriptomes and metabolomes of Pyropia yezoensis conchocelis unveil the roles of pyPGK, pyBCKDHA, and pyDLD in response to freshwater soaking. Int. J. Biol. Macromol. 285, 138193. https://doi.org/10.1016/j.ijbiomac.2024.138193.
10.1016/j.ijbiomac.2024.138193Hwang MS, Kim SM, Ha DS, Baek JM, Kim HS and Choi HG. 2005. DNA sequences and identification of Porphyra cultivated by natural seeding on the southwest coast of Korea. Algae 20, 183-196. https://doi.org/10.4490/ALGAE.2005.20.3.183.
10.4490/ALGAE.2005.20.3.183Hwang MS, Kim SO, Lee YS, Park EJ, Kim SC, Ha DS, Gong YG, Baek JM and Choi HG. 2010. Isolation and characterization of pure lines of pigmentation and morphological mutants in Porphyra tenera Kjellman (Bangiales, Rhodophyta). Kor. J. Fish. Aquat. Sci. 43, 495-502. https://doi.org/10.5657/kfas.2010.43.5.495.
10.5657/kfas.2010.43.5.495Jiang H, Ding H, Zhang P, Wang T and Yan X. 2020. Selection and characterization of an improved strain (A-13) of Pyropia yezoensis (Bangiales, Rhodophyta). Aquat. Bot. 163, 103213. https://doi.org/10.1016/j.aquabot.2020.103213.
10.1016/j.aquabot.2020.103213Kim HS, Choi HG, Hwang MS, Jeon YJ, Yarish C and Kim JK. 2022. Concise review of the genus Neopyropia (Rhodophyta: Bangiales). J. Appl. Phycol. 34, 1805-1824. https://doi.org/10.1007/s10811-022-02776-1.
10.1007/s10811-022-02776-1Kim JH, Choi SJ and Lee S. 2019. Effects of temperature and light on photosynthesis and growth of red alga Pyropia dentata (Bangiales, Rhodophyta) in a conchocelis phase. Aquaculture 505, 167-172. https://doi.org/10.1016/j.aquaculture.2019.02.058.
10.1016/j.aquaculture.2019.02.058Kim NG and Notoya M. 2004. Life history of Porphyra seriata Kjellman (Bangiales, Rhodophyta) from Korea in laboratory culture. Algae 19, 303-309. https://doi.org/10.4490/ALGAE.2004.19.4.303.
10.4490/ALGAE.2004.19.4.303Kim NG. 1999. Culture studies of Porphyra dentata and P. pseudolinearis (Bangiales, Rhodophyta), two dioecious species from Korea. Hydrobiologia 398, 127-135. https://doi.org/10.1007/978-94-011-4449-0_15.
10.1007/978-94-011-4449-0_15Kim NG. 2001. Physiological study on the hybrid by interspecific crossing between Porphyra pseudolinearis and P. dentata (Bangiales, Rhodophyta), two dioecious species in culture. J. Aquac. 13, 353-357.
Lee HJ, Park EJ and Choi J. 2019. Isolation, morphological characteristics and proteomic profile analysis of thermos-tolerant Pyropia yezoensis mutant in response to high-temperature stress. Ocean Sci. J. 54, 65-78. https://doi.org/10.1007/s12601-018-0060-9.
10.1007/s12601-018-0060-9Li X, Yang L and He PM. 2011. Formation and growth of free-living conchosporangia of Porphyra yezoensis: effects of photoperiod, temperature and light intensity. Aquac. Res. 42, 1079-1086. https://doi.org/10.1111/j.1365-2109.2010.02691.x.
10.1111/j.1365-2109.2010.02691.xLiang Z, Liu F, Wang W, Zhang P, Yuan Y, Liu Y, Yao H, Jia R, Sun X and Wang F. 2022. Physiological and biochemical responses to light and temperature stress in free-living conchocelis of Neopyropia katadae (Bangiales, Rhodophyta). J. Appl. Phycol. 34, 1059-1072. https://doi.org/10.1007/s10811-022-02691-5.
10.1007/s10811-022-02691-5Lin Y, Xu K, Xu Y, Ji D, Chen C, Wang W and Xie C. 2021. Transcriptome co-expression network analysis identifies key genes regulating conchosporangia maturation of Pyropia haitanensis. Front. Genet. 12, 680120. https://doi.org/10.3389/fgene.2021.680120.
10.3389/fgene.2021.68012034276783PMC8278576Liu HL, Shuai L, Duan DL and Xu HS. 2002. Axenic culture of free-living conchocelis of Porphyra yezoensis and Porphyra haitanensis. Chin. J. Ocean. Limnol. 20, 62-66. https://doi.org/10.1007/BF02846612.
10.1007/BF02846612Lu S and Yarish C. 2011. Interaction of photoperiod and temperature in the development of conchocelis of Porphyra purpurea (Rhodophyta: Bangiales). J. Appl. Phycol. 23, 89-96. https://doi.org/10.1007/s10811-010-9541-7.
10.1007/s10811-010-9541-7Luo Q, Zhu Z, Zhu Z, Yang R, Qian F, Chen H and Yan X. 2014. Different responses to heat shock stress revealed heteromorphic adaptation strategy of Pyropia haitanensis (Bangiales, Rhodophyta). PLOS ONE 9, e94354. https://doi.org/10.1371/journal.pone.0094354.
10.1371/journal.pone.009435424709783PMC3978056Ma M, Liu Q, Huang L, Zhang P, Chen H, Chen J, Luo Q, Wang T and Yang R. 2023. Purification of conchocelis of Neoporphyra haitanensis by the method of dominant Bacteria enrichment comprehensive treatment. Aquaculture 573, 739594. https://doi.org/10.1016/j.aquaculture.2023.739594.
10.1016/j.aquaculture.2023.739594McLachlan J. 1973. Growth media-marine. In: Handbook of phycological methods. Culture methods and growth measurements. Stein JR, ed. Cambridge University Press, Cambridge, 25-51.
Ott FD. 1965. Synthetic media and techniques for the xenic cultivation of marine algae and flagellate. Va. J. Sci. 16, 205-218.
Park CS and Hwang EK. 2014. Isolation and evaluation of a strain of Pyropia yezoensis (Bangiales, Rhodophyta) resistant to red rot disease. J. Appl. Phycol. 26, 811-817. https://doi.org/10.1007/s10811-013-0183-4.
10.1007/s10811-013-0183-4Park CW, Choi KJ, Soh EH and Koh HJ. 2016. Study on the future development direction of plant variety protection system in Korea. Korean J. Breed. Sci. 48, 11-21. https://doi.org/10.9787/KJBS.2016.48.1.011.
10.9787/KJBS.2016.48.1.011Park EJ. 2023. Cross-breeding of Neopyropia spp. (Bangiales, Rhodophyta) using CAPS (Cleaved Amplified Polymorphic Sequence) markers. Korean J. Fish. Aquat. Sci. 56, 124-132. https://doi.org/10.5657/KFAS.2023.0124.
10.5657/KFAS.2023.0124Provasoli L. 1968. Media and prospects for the cultivation of marine algae. In: Cultures and Collections of Algae: Proceedings of the U.S.-Japan Conference, Hakone, September 1966. Watanabe H and Hattori A, eds. Japanese Society of Plant Physiology, Hakone, 63-75.
Sasuga K, Yamanashi T, Nakayama S, Ono S and Mikami K. 2017. Optimization of yield and quality of agar polysaccharide isolated from the marine red macroalga Pyropia yezoensis. Algal Res. 26, 123-130. https://doi.org/10.1016/j.algal.2017.07.010.
10.1016/j.algal.2017.07.010Seo HM, Han LY, Lee WJ, Im HJ and Park EJ. 2025. Effects of temperature, irradiance and nutrients on the growth and maturation of conchocelis in Pyropia yezoensis and Pyropia dentata. Korean J. Fish. Aquat. Sci. 58, 445-452. https://doi.org/10.5657/KFAS.2025.0445.
10.5657/KFAS.2025.0445Shin YJ, Min SR, Kang DY, Lim JM, Park EJ, Hwang MS, Choi DW, Ahn JW, Park YI and Jeong WJ. 2018. Characterization of high temperature-tolerant strains of Pyropia yezoensis. Plant Biotechnol. Rep. 12, 365-373. https://doi.org/10.1007/s11816-018-0499-2.
10.1007/s11816-018-0499-2Wang D, Yu X, Xu K, Bi G, Cao M, Zelzion E, Fu C, Sun P, Liu Y, Kong F, Du G, Tang X, Yang J, Tang L, Zhao Y, Ge Y, Zhuang Y, Mo Z, Chen Y, Gao T, Guan X, Chen R, Qu W, Sun B, Bhattacharya D and Mao Y. 2020. Pyropia yezoensis genome reveals diverse mechanisms of carbon acquisition in the intertidal environment. Nat. Commun. 11, 4028. https://doi.org/10.1038/s41467-020-17689-1.
10.1038/s41467-020-17689-132788591PMC7423979Yan N, Ding HC and Yan XH. 2024. Selection and characterization a new strain (PS-M4) of Pyropia suborbiculata with fast growth based on 60Co-γ ray irradiation. J. Appl. Phycol. 36, 2769-2780. https://doi.org/10.1007/s10811-024-03268-0.
10.1007/s10811-024-03268-0Zhong ZH, Wang WJ, Sun XT, Liu FI, Liang ZR, Wang FJ and Chen WZ. 2016. Developmental and physiological properties of Pyropia dentata (Bangiales, Rhodophyta) conchocelis in culture. J. Appl. Phycol. 28, 3435-3445. https://doi.org/10.1007/s10811-016-0877-5.
10.1007/s10811-016-0877-5

