

1. 서론
2. 재료 및 방법
2.1 직립지와 포복지 생장 패턴
2.2 염분과 영양염 농도에 따른 생장
2.3 통기기간과 생장
2.4 통계분석
3. 결과
3.1 직립지와 포복지 생장 패턴
3.2 염분과 영양염 농도에 따른 생장
3.3 통기 기간의 영향
4. 고찰
1. 서론
옥덩굴 속 해조류는 열대에서 아열대 해역까지 분포하는 관상(siphonous) 녹조류로서 전 세계적으로 400여종이 기재되었고 현재 105종이 인정되며, 그 중에서 15종이 식용으로 활용된다(de Gaillande et al. 2017; Guiry and Guiry 2021). 바다포도(sea grape)로 불리는 열대해역의 대표적인 식용종은 Caulerpa lentillifera와 C. racemosa var. turbinata이며, C. lentillifera는 일본 오키나와(Okinawa)에서 관광객에게 특산품으로 인기가 높은 것으로 알려져 있다(Ratana-arpon and Chirapart 2006; Shi 2008). 특히, C. lentillifera는 부드러운 식감과 고도불포화지방산(polyunsaturated fatty acids, PUFA)과 다중필수아미노산(multiple essential amino acids)을 다량 함유하지만, 지질 함량이 낮아 슈퍼푸드(superfood)로 자리매김하여(Niwano et al. 2009; Saito et al. 2010), 필리핀, 베트남, 일본, 타이완 등에서 양식되고 있다(Kurashima et al. 2003; Matanjun et al. 2009; de Gaillande et al. 2017).
옥덩굴 속의 식용 해조류는 엽체 절편으로 생장하는 클론 해조류(clonal seaweed)로서 김, 미역, 다시마와 같이 채묘와 종묘 배양 등의 복잡한 단계(multistep)를 가진 비클론 해조류(non-clonal seaweed)에 비해 노동력이 적게 드는 장점이 있다(Bast 2013). 하지만, 클론 해조류는 엽체별 생장속도가 다르므로 생산량 증대를 위해서는 클론내 변이체(intra-clonal variant) 연구를 통하여 생장이 우수한 양식 엽체 선별(strain selection) 과정이 요구된다(Santelices 2001). 양식 해조류의 생산량 증대를 위한 최적 온도, 광도, 염분 및 영양염 등의 환경조건 탐색에 대한 생리적 연구가 필요한 것처럼(Horstmann 1983; Guo et al. 2015a, b), 옥덩굴 속 해조류 (Caulerpa taxifolia, C. scalpelliformis, C. prolifera, C. lentillifera) 생장을 위한 최적 환경 조건 탐색에 대한 연구도 수행되었다(O’Neal and Prince 1988; Komatsu et al. 1997; Ukabi et al. 2013; Guo et al. 2015a, b). 한 예로, C. lentillifera의 생장을 위한 최적 염분은 태국산이 25-30 psu였으며(Deraxbudsarakom et al. 2003), 베트남산과 일본산이 각기 36 psu와 35 psu로서(Wang 2011; Guo et al. 2015b), 동일 종임에도 서식 해역에 따라 염분 요구성이 다름을 확인하였다. 또한, C. lentillifera의 생장을 위한 적정 P와 N의 농도는 0.1 mmol L−1 (PO4-P)와 0.5 mmol L−1 (NO3-N)로 확인되었으며(Guo et al. 2015a), N:P가 8:1의 비율에서, 그리고 0.6 mmol L−1 (NO3-N)에서 최대 생장이 기록되어(Deraxbudsarkom et al. 2003), N과 P의 농도와 이들의 비율이 생장에 영향을 미치는 것으로 나타났다.
우리나라에서 아열대지표종인 옥덩굴(Caulerpa okamurae Weber-van Bosse)의 지리적 분포와 개체군 생장에 대한 시·공간적 변동은 몇몇 해조학자들에 의해 연구가 이루어졌다(Choi 2007; Park et al. 2014; Kim et al. 2016). 현재, 옥덩굴은 제주도, 남해안, 울릉도와 독도 등에 분포하고 있으며, 조하대에 서식하는 것으로 알려져 있다(Choi et al. 2000; Choi 2007; Park and Choi 2009; Park et al. 2014; Kim et al. 2016). 국내산 옥덩굴은 건중량 100 g에 포함된 단백질이 12.8 g으로 지방(0.3 g)에 비해 훨씬 많았고, 탄수화물(31.2 g)과 식이섬유(28.4 g)도 높았으나 중금속(Hg, As, Cd, Pb) 함량은 정량한계(LOQ, limit of quantitation) 이하로 확인되었다. 온도와 광도가 옥덩굴의 생장 및 성숙에 미치는 영향에 대한 실내 배양 연구가 시도되었으며(Choi et al. 2000; Hwang et al. 2003), 최근에 Gao et al. (2019)은 옥덩굴 직립지와 포복지가 광염성(20~45 psu) 생장을 보였으며, 최적 염분 농도는 30 psu로 기록하였다. 하지만, 현재까지 염분과 영양염의 조합조건에서 옥덩굴 직립지와 포복지 생장과 광합성에 미치는 영향은 확인되지 않았다. 따라서, 본 연구는 옥덩굴 육상양식의 기초자료를 확보하기 위하여, 직립지와 포복지의 생장 패턴과 염분과 영양염 농도의 조합, 그리고 영양염 순환과 관련된 일별 통기 기간이 엽체 생장에 미치는 영향을 확인하기 위하여 수행되었다.
2. 재료 및 방법
2.1 직립지와 포복지 생장 패턴
옥덩굴 엽체는 전남 진도군(34°39' N, 126°27' E)의 조간대 하부 암반에서 2019년 6월 5일에 채집하였으며 현장 해수와 함께 플라스틱 지퍼백에 넣고 아이스박스에 담아 실험실로 운반하였다. 엽체 표면의 착생 동∙식물과 불순물은 핀셋과 유화용 붓으로 제거한 후, 여과 및 멸균 해수로 수회 세척하였다. 실험용 옥덩굴 직립지와 포복지를 길이 3cm와 5cm로 절단하여 멸균해수가 담긴 plant culture dish에 넣고, 상처 회복을 위하여 20℃, 20 µmol photons m-2 s-1와 16L:8D (Light: Dark)의 광주기로 설정된 항온실에서 정치배양하였다.
배양 1일 후, 배양액 200 mL이 담긴 plant culture dish 6개를 준비하였고, 그 중에서 3개 배양용기에는 직립지, 다른 3개 용기에는 포복지 3개체씩을 넣었다. 배양액은 PES30 (30% 희석한 PES, Provasoli 1968)을 사용하였고 25℃, 40 µmol photons m-2 s-1와 16L:8D로 세팅된 인큐베이터에서 10일 동안 배양하였다. 배양액은 4일 간격으로 전량 교환하였으며, 매일 한 개체를 무작위로 골라 해부현미경하에서 사진 촬영하였다.
2.2 염분과 영양염 농도에 따른 생장
염분과 영양염 농도의 상호작용이 옥덩굴 생장 및 엽록소 형성에 미치는 영향을 파악하기 위하여, 직립지 81개체와 포복지 81개체를 실험 2일 전에 절단하여 정치 배양하였다. 직립지와 포복지는 다양한 염분(20, 30, 40 psu)과 영양염 농도(ASW, PES30, PES50)의 조합조건인 9개 실험구(salinity 3×nutrients 3)로 구분하였고 실험구별 반복구 3개씩을 두었다. 저영양염 농도(low nutrients)는 멸균해수(ASW, autoclaved seawater), 중영양염 농도(mid nutrients)는 PES30, 그리고 고영양염 농도(high nutrients)는 PES50으로 표기하였다. 옥덩굴 직립지(길이 3 cm)와 포복지(길이 3 cm)는 200 mL 배양액이 담긴 27개의 plant culture dish에 3개체씩 배분하였다. 배양액에 GeO2 (5 mg L-1)을 첨가하여 돌말류(diatoms) 생장을 억제시켰으며, 배양액은 4일 간격으로 전량 교환하면서 20일 동안 실험을 실시하였다. 실험용 염분 농도는 멸균해수(30 psu)에 증류수 첨가(20 psu) 혹은 천일염 첨가(40 psu)로 조제하였으며, 염분계(Atago, S/Mill, Japan)로 농도를 측정하여 확인하였다. 배양 20일 후, 엽체 무게(g wet weight)는 전자저울로 측정하였고 상대생장률(RGR, relative growth rate)은 배양 개시와 종료 시 무게로 계산하였다(Choi et al. 2008).
RGR (% day-1) = 100 ln (Wt / W0) / T
Wt는 배양 종료 시 무게, W0은 배양 개시 때의 무게, T는 배양일 수(day)를 나타낸다.
엽록소 함량은 염분과 영양염 농도별로 옥덩굴 엽체의 무게를 측정한 후, 각 실험구에서 무작위로 선택된 엽체 5 g (습중량)을 sea sand (2 g)와 함께 막자 사발에 넣어 분쇄하였다. 냉장고(4℃)에 보관된 90% 아세톤 6 mL을 막자사발에 넣고 4분 동안 엽록소를 추출하였으며, 이 용액을 test tube (15 mL)에 담은 뒤 이동시켰으며, 다시 막자 사발에 아세톤(4 mL)을 넣고 3분 동안 반복 추출하여 동일한 test tube에 첨가하였다. 아세톤 추출액을 원심 분리한 후 상층액을 흡광도 630, 645, 647, 663, 664 nm에서 분광광도계(spectrophotometer)로 측정하였다(Haryatfrehni et al. 2015). 엽록소 총량과 chl a의 함량은 아래의 식과 같이 계산하였다.
Total chlorophyll (mg L−1) = 8.02A663 + 20.21A645 (Arnon 1949)
Chlorophyll a (mg L−1) = 11.85A664 – 1.54A647 – 0.08A630 (Jeffrey and Humphrey 1975)
2.3 통기기간과 생장
옥덩굴 생장에 일별 통기 기간(aeration period)이 미치는 영향을 파악하기 위하여, 2 cm 미만의 직립지가 1~2개 붙어 있는 포복지를 준비하였다. 2일 후, 800 mL의 PES30 배양액이 담긴 비이커 15개에 포복지 2 g (in wet weight)씩을 넣었다. 옥덩굴 포복지를 하루 정치 배양 후, 다양한 일별 통기 기간(0, 8, 12, 16, 24 h day-1)로 3개 비이커(반복구)를 배정하여 15일 동안 배양하였다. 통기 기간은 에어펌프에 연결된 타이머로 설정하였다. 옥덩굴은 20℃, 20 µmol photons m-2 s-1, 16L:8D의 동일한 환경에서 배양하였으며, 배양액은 3일 간격으로 전량 환수하였다.
2.4 통계분석
옥덩굴의 생장에 통기 기간이 미치는 영향은 one-way ANOVA test로, 그리고 염분·영양염 농도의 조합의 영향은 two-way ANOVA test로 유의차 검정을 실시하였다. 평균에 대한 유의차가 발견되면 Tukey’s HSD test 방법으로 사후 검정을 실시하였으며(Sokal and Rohlf 1995), 데이터 분석을 위해 사용한 통계 프로그램은 STATISTICA version 10.0이었다.
3. 결과
3.1 직립지와 포복지 생장 패턴
옥덩굴 직립지와 포복지의 생장실험에서 직립지 절단면의 상처는 3~4일 후에 완전하게 치료되었으며, 배양 5일 후에 새로운 포복지와 가근(rhizoid)이 자랐고 10일 후에는 수많은 가근과 포복지가 관찰되었다(Fig. 1A-D). 포복지는 절단면에서 약간의 가근 형성과 포복지 생장이 관찰되었고 5일 후에는 새롭게 생장한 포복지에서 직립지와 가근이 발달하였다. 배양 10일 후에는 최초의 포복지에서 새롭게 형성된 여러 개의 직립지가 관찰되었다(Fig. 1E-H).

3.2 염분과 영양염 농도에 따른 생장
염분과 영양염 농도의 조합 조건에서 20일 배양한 결과, 옥덩굴 포복지(RGR; 0.71~3.52% day-1)가 직립지(-0.57~2.08% day-1)에 비해 빠른 생장을 보였다(Fig. 2A, B). 옥덩굴 직립지와 포복지의 최대 생장은 30 psu에서 일어났고 최소 생장은 20 psu에서 관찰되었다. 또한, 영양염 농도별 옥덩굴 생장은 직립지에서는 중영양인 PES30에서, 그리고 포복지에서는 고영양 농도인 PES50에서 최대였다. 옥덩굴 직립지의 상대생장률은 염분별 유의차만 존재하였으며(P <0.001, Table 1), 포복지의 생장은 염분과 영양염 농도별 유의차가 나타났고 염분·영양염 농도의 상호작용에도 영향을 받는 것으로 확인되었다(P <0.05, Table 1).
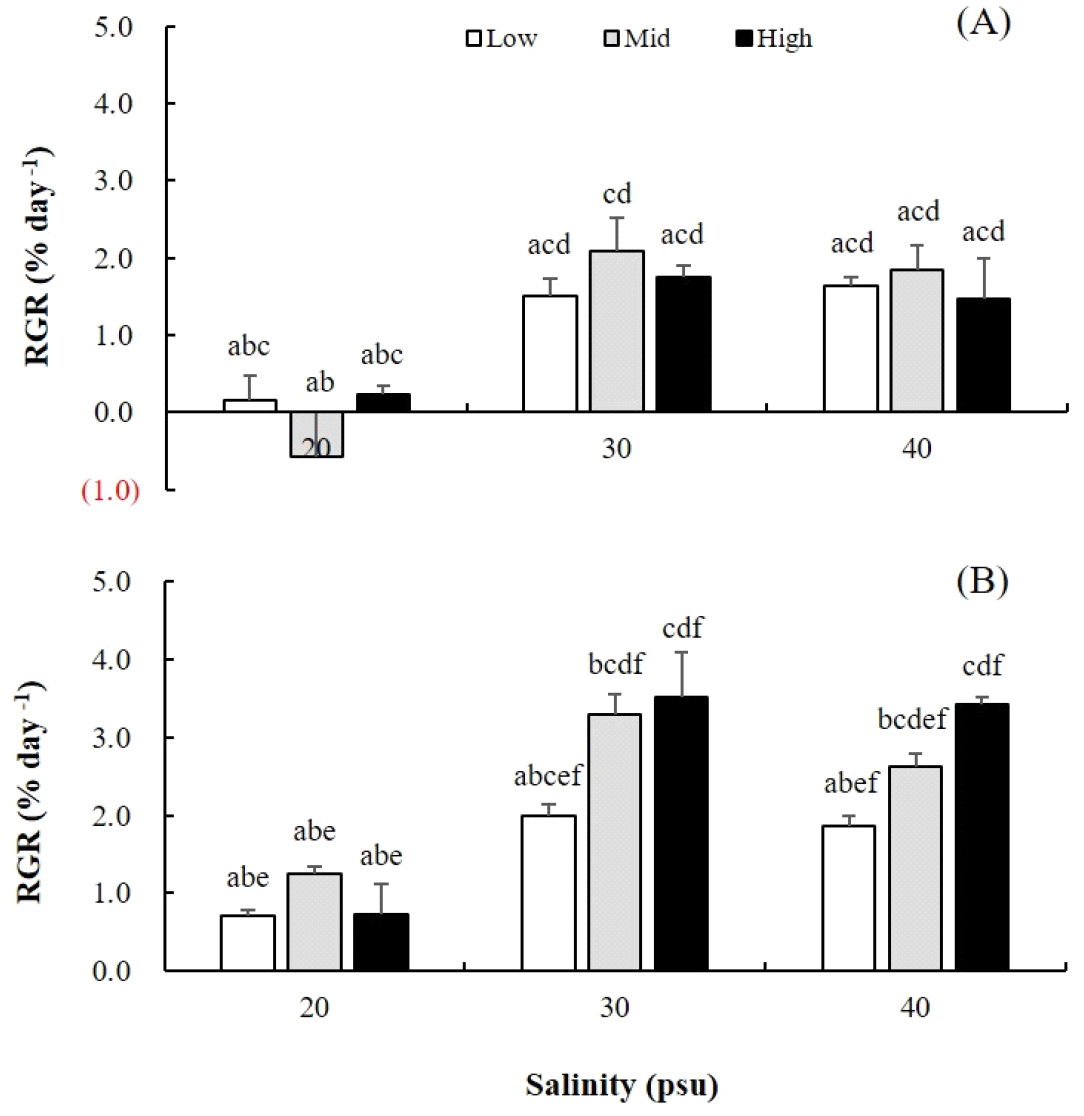
Fig. 2
RGRs of erect fronds (A) and stolons (B) of Caulerpa okamurae. They were cultured for 20 days under three salinities (20, 30, 40 psu) and three nutrient levels (Low, Mid, High). Low, mid, high present autoclaved seawater, PES30, and PES50 (50% dilution of PES), respectively. Culture conditions were 20°C, 20 µmol photons m-2 s-1, 16:8 h L:D. Vertical bars indicate mean±SE (n=3 replicates).
Table 1.
Analysis of variance (ANOVA) for the effects of salinities and nutrient levels on RGR and morphological parameters of erect frond and stolon frond of Caulerpa okamurae.
옥덩굴 직립지와 포복지는 20 psu의 낮은 염분 농도에서는 생장 지연되었으며, 직립지에서는 말단가지(ramuli)가 탈락되기도 하였다. 염분 농도 30 psu와 40 psu에서 배양한 옥덩굴 직립지에서는 새로운 포복지가 많이 형성되었으며 이 포복지로부터 짧은 직립지가 1~2개 형성되었다. 한편, 옥덩굴 포복지 실험구에서는 최초의 포복지에서 새로운 직립지가 2~4개 형성되었다. 영양염 농도별 옥덩굴의 생장 반응을 보면, 직립지와 포복지는 멸균 해수(저영양염)에서는 포복지 생장만 보였으며, 다른 영양염 농도에 비해 연한 초록색을 보였다. 직립지와 포복지는 배양액의 영양염 농도가 증가될수록 엽체의 색상은 진한 녹색을 나타냈고, 고염분·고영양 농도 실험구에서는 배양 기간이 길어질수록 남조류와 돌말류 등 착생조류의 감염이 많았다.
배양 20일 후, 엽록소 총량은 옥덩굴 직립지에서 0.47~1.09 mg g-1이었고, 포복지에서 0.22~0.61 mg g-1로 직립지가 1.7배 높았다(Fig. 3A, B). 직립지와 포복지 엽록소 총량은 저영양인 멸균해수에서 최소였고 PES30과 PES50에서 높게 나타났다. 특히, 옥덩굴 직립지의 엽록소 총량은 염분과 관련성이 높게 나타났으며, 저염분(20 psu)과 고염분(40 psu)에서는 중영양 농도인 PES30에서 최대였으며, 생장이 최대로 일어나는 30 psu에서는 고영양염 상태인 PES50에서 빠른 생장을 보였다(Fig. 3A). 옥덩굴 포복지의 엽록소 총량은 모든 염분(20, 30, 40 psu)에서 공통적으로 멸균해수에서 최소였고 PES30과 PES50에서 높은 값을 보였으며, 중영양과 고영양염에서 엽록소 총량에 유의한 차이는 없었다(Fig. 3B).
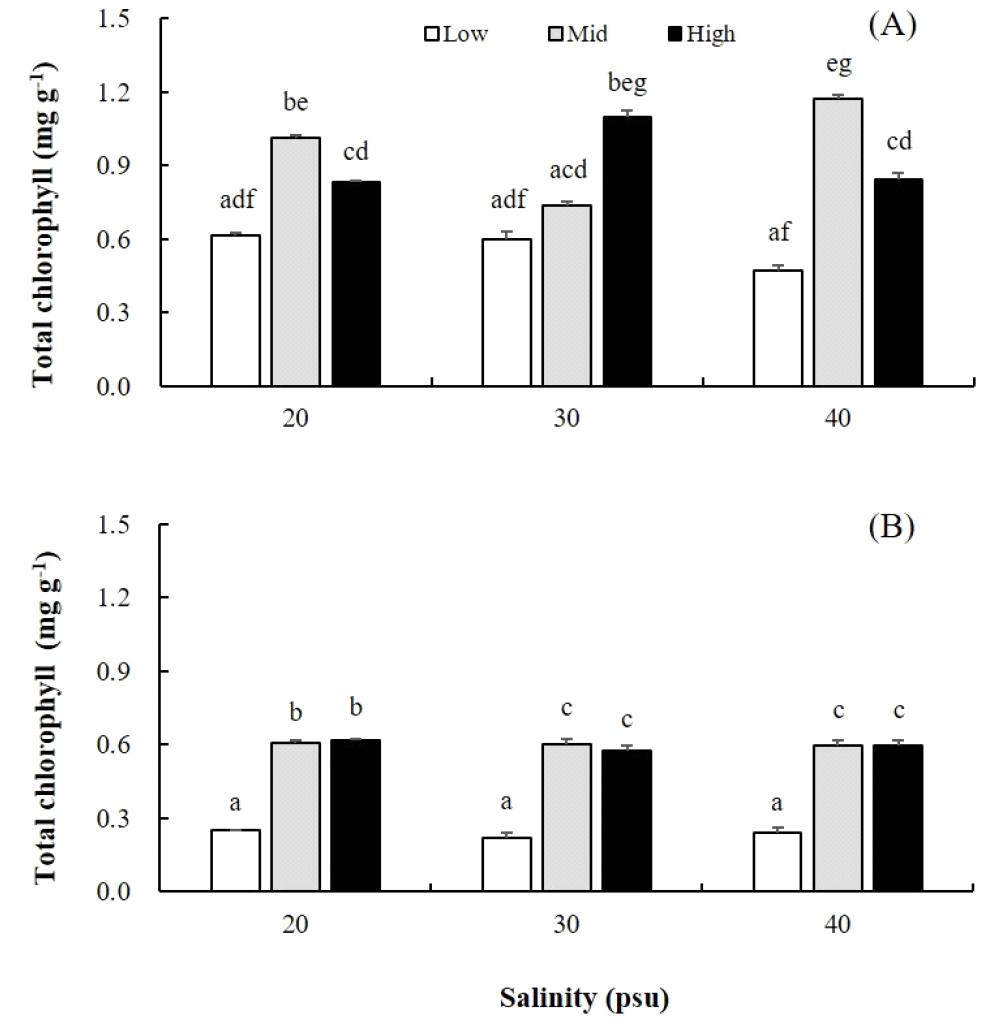
Fig. 3
Effects of salinities and nutrient levels on the contents of total chlorophyll of erect frond (A) and stolon (B) of Caulerpa okamurae after 20 days in culture. They were cultured under three salinities (20, 30, 40 psu) and three nutrient levels (Low, Mid, High). Low, Mid, High present autoclaved seawater, PES30, and PES50 (50% dilution of PES), respectively. Culture conditions were 20°C, 20 µmol photons m-2 s-1, 16L:8D. Vertical bars indicate mean±SE (n=3 replicates).
엽록소 a 함량은 옥덩굴 직립지에서 포복지에 비해 1.6배 높게 나타났으며, 모두 저영양에서 최소(직립지, 0.26; 포복지, 0.12 mg g-1)였다. 직립지의 엽록소 a 함량은 중영양에서 0.74 mg g-1으로, 그리고 포복지는 고영양에서 0.42 mg g-1으로 최대였다(Fig. 4A, B). 옥덩굴 직립지에서 엽록소 총량과 chl a의 함량은 염분별 유의차를 보였으며, 염분·영양염 농도의 상호작용도 확인되었다(Table 1).
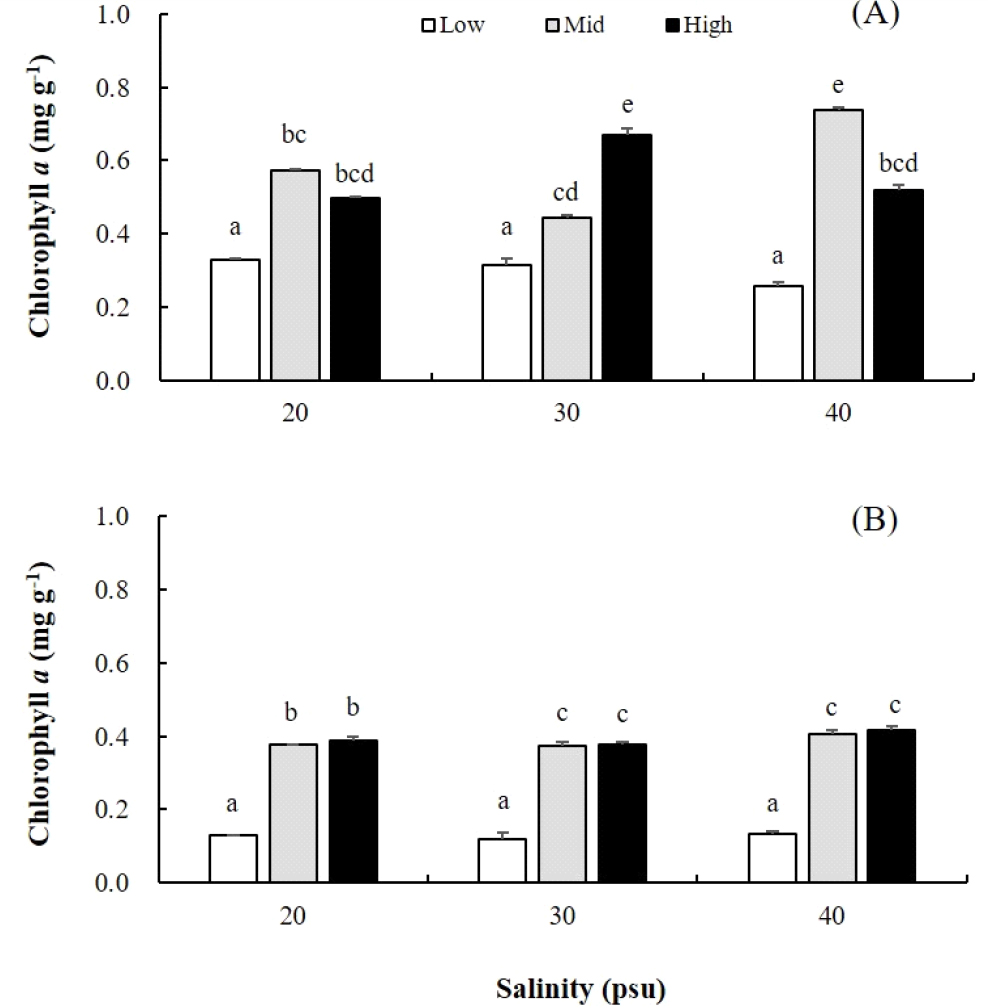
Fig. 4
Effects of salinities and nutrient levels on the contents of chlorophyll a of erect fronds (A)and stolons (B) of Caulerpa okamurae after 20 days. They were cultured under three salinities (20, 30, 40 psu) and three nutrient levels (Low, Mid, High). Low, Mid, High present autoclaved seawater, PES30, and PES50 (50% dilution of PES), respectively. Culture conditions were 20°C, 20 µmol photons m-2 s-1, 16L:8D. Vertical bars indicate mean±SE (n=3 replicates).
3.3 통기 기간의 영향
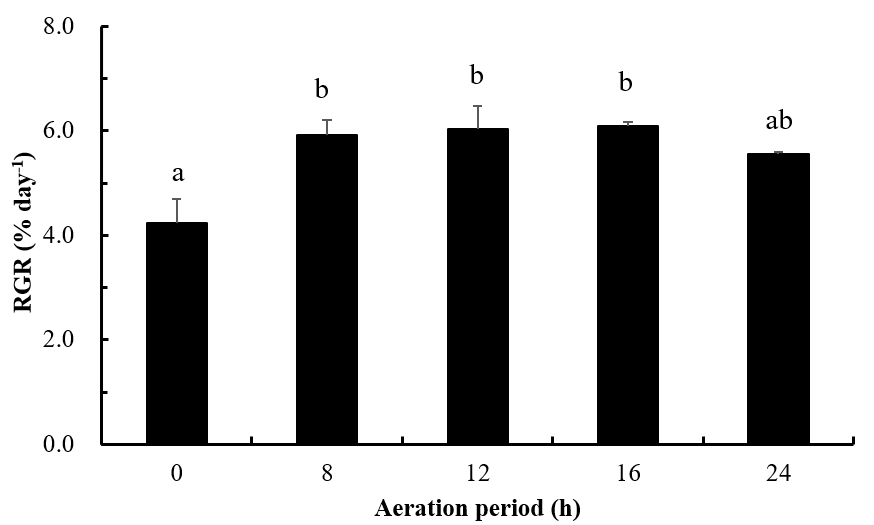
옥덩굴 포복지는 최초에 2 g (in wet weight)을 배양 용기에 넣고 15일 동안 다양한 일별 통기기간 조건에서 배양한 결과 3.85~5.15 g으로 생장하였다. 포복지 상대생장률은 정치 배양(no aeration)에서 4.24% day-1로 최소였고, 16 h 통기배양에서 6.10% day-1로 최대였고, 정치배양에 비해 16 h에서 1.4배 빠른 생장을 보였다(Fig. 5). 포복지 생장은 정치 배양(0 h)에 비해 통기 배양(8, 12, 16 h day-1)에서 높게 나타났으나(Tukey’s HSD test, F4,10=16.08, P<0.01), 정치 배양과 연속 통기 배양(24 h day-1)의 엽체 생장은 유의차를 보이지 않았다(P>0.05, Fig. 4).
4. 고찰
옥덩굴 류(Caulerpa spp.) 양식은 엽체 절편을 이용한 영양체 번식 방법(vegetative propagation method)으로 수행되고 있다(Smith and Walters 1999; Phillips 2009; Zubia et al. 2020). 옥덩굴(Cauelrpa okamurae)의 유성생식은 실내 배양과 야외 개체군에서 일부 관찰되었으나(Hwang et al. 2003), 종묘 생산에는 활용되지 않았다. 대신, 옥덩굴 엽체의 절편은 실내 배양과 야외 양식에서 빠른 생장을 보였으며(Choi et al. 2000; Gao et al. 2019), 최근에 직립지, 포복지, 혹은 직립지+포복지의 생장 실험에서 포복지의 생장률이 4.93% day-1로 최대였다(Shin et al. 2021). 옥덩굴 부위별 생장 및 환경에 대한 내성 차이는 밝혀졌으며(Gao et al. 2019), 온대해역에서 수온이 낮은 겨울철에 포복지로 살아가는 아열대지표종의 생존전략과 관련이 있는 것으로 판단된다. Cauelrpa okamurae taxifolia와 C. cylindracea는 포복지 생장과 엽체 절편에 의한 무성생식 방법으로 지리적 분포 범위를 확대하는 생태학적 침입종으로 잘 알려져 있다(Komatsu et al. 1997; Wright and Davis 2006; Zubia et al. 2020). 따라서, 수온상승과 해양산성화 같은 해양의 기후 변화와 관련하여 아열대 지표종 옥덩굴의 절편에 의한 지리적 분포 확대와 절편을 이용한 대량 양식 방법을 개발하는데 관심을 가져야 할 것으로 판단된다.
옥덩굴 엽체를 다양한 염분(20, 30, 40 psu)과 영양염 농도(low, mid, high)의 조합으로 배양하였을 때, 염분 30 psu (상대생장률, 1.78% day-1)와 40 psu (1.64% day-1)에서 빠른 생장을 보였다. 엽록소 총량(0.83 mg g-1)과 엽록소 a의 함량(0.50 mg g-1)은 염분 40 psu에서 최대였으나, 20~30 psu와 유의차는 없었다. 옥덩굴 직립지를 15~45 psu에서 배양하였을 때, 상대생장률은 5.91% day-1로 30 psu에서 최대였으며(Gao et al. 2019), C. lentillifera 직립지의 염분 농도별(15~55 psu) 실험에서 최대 생장은 35 psu (상대생장률, 2.04% day-1)에서 기록되었다(Guo et al. 2015b). 옥덩굴 류는 종별 생장을 위한 최적 염분 농도는 다른 것으로 확인되었으며(Guo et al. 2015b; Gao et al. 2019), 동일한 종일지라도 배양조건과 배양시기에 따라 생장 차이를 보인다는 것을 확인하였다.
본 연구에서 옥덩굴의 포복지 절편은 정치배양(0 h day-1)에 비해 통기배양에서 생장은 촉진되었으나, 흥미롭게도 연속 통기배양(24 h day-1)에 비해 통기 기간이 8~16 h day-1일 때 생장이 좋은 것으로 확인되었다. 꼬시래기 류(Gracilaria sp. Strain G-16)의 통기기간별(4, 8, 12, 16, 20, 24 h day-1) 배양실험에서, 생물량은 4 h day-1에서는 감소하였지만, 다른 조건에서는 증가되는 것으로 확인되어(Guerin and Bird 1987), 본 연구 결과와 같이 통기(aeration)가 해조류 생장을 촉진하는 것으로 나타났다. 하지만, 옥덩굴 포복지 절편이 8~16 h day-1의 통기 기간에서 연속 통기(24 h day-1) 조건에 비해 빠른 생장을 나타낸 것은 통기로 인해 발생한 공기방울(bubble)이 엽체에 도달하는 광량을 감소시켰기 때문으로판단된다. 또한, 옥덩굴 직립지와 기질 부착용 가근을 만드는 포복지 절편의 생장이 통기 기간에 따라 다르게 나타나는지에 대한 추가적인 연구가 요구된다. 본 연구 결과, 통기배양에서 옥덩굴의 빠른 생장은 해수 흐름으로 인해 광합성에 필요한 CO2와 영양분 공급이 증가되었기 때문으로 판단된다(Guerin and Bird 1987; Flagella et al. 2008; Msuya and Neori 2008).
옥덩굴 류는 광범위한 온도, 광도와 염분 농도에서 생장이 가능하다고 알려져 있다(Komatsu et al. 1997; Friedlander et al. 2006; Ukabi et al. 2013; Guo et al. 2015a, b; Gao et al. 2019). 본 연구 결과, 옥덩굴 엽체의 절편 생장은 다양한 염분과 영양염 농도에서 일어났으며, 직립지에 비해 포복지가 빠른 것으로 확인되었고, 최적 생장은 30~40 psu의 염분, PES30와 PES50의 영양염 농도에서 일어났다. 이외에도, 옥덩굴 포복지 생장은 일별 통기 기간에 영향을 받았으며, 생장을 위한 최적 통기 기간은 8~16 h day-1로 확인되었다.
