

1. 서론
한국의 연안 저서 암반 생태계에서 유용한 엽상해조의 소실과 광범위한 갯녹음현상(barren ground)의 확산은 세계적인 기후변화와 맞물려 해양생태계 변화를 극적으로 보여주는 현상으로 나타났다. 엽상해조(foliose algae)의 소실로인해 무절산호조류(crustose coralline algae)가 노출되면서 갯녹음현상(barren ground)의 원인생물로 간주되어 주목을 받는 계기가 되었다(MOF 2002; NFRDI 2008).
무절산호조류(CCA)는 전 세계 해양에 널리 분포하며 암반기질 전체를 피복하기도 한다(Keats and Maneveldt 1994; Maneveldt et al. 2007). 몸 전체가 석회화되어 있으며, 지붕이 있는 형태의 생식세포 구조를 갖고, 영양세포들은 섬유상으로 체계화되어있다(Johansen 1981; Harvey et al. 2005). 주요한 1차 생산자로써 12.8~22.8 mmol m-2h-1의 산소를 생산하며(Chisholm 2003), 얼음으로 뒤덮인 극지방의 낮은 광량조건인 저서환경에서도 서식하는 등, 1차 생산자로서의 역할을 하고 있다(Roberts et al. 2002). 잠수정을 이용한 조사를 통해 표층의 0.001% 빛만이 도달되는 수심 200m 지점에서도 무절산호조류들이 군락을 이루어 서식하는 것이 밝혀졌다(Litter et al. 1986). 또한, 산호초(coral reef) 생태계에서 산호(coral) 스스로는 거대한 조초를 유지할 수 없고, 무절산호조류와 함께 생육하여야만 그 골격을 유지할 수 있다(Woelkerling 1988). 무절산호조류는 1차 생산자로서 그리고 해양생태계를 유지하는 하나의 구성원으로 중요한 위치에 자리하고 있다.
해양환경에서 중요한 위치에 있는 무절산호조류에 대한 분류학적, 생태적인 연구가 진행되고 있지만(Borowitzka and Townsend 2001; Broadwater et al. 2002; Harvey et al. 2003; Adey et al. 2005; Maneveldt et al. 2007; Broom et al. 2008; Maneveldt et al. 2008; Caragnano et al. 2009; Francesca et al. 2011), 배양에 관한 연구는 매우 미흡하게 진행된 상태이고, 동일한 종(species)일지라도 서식환경에 따라 생장과 생식에 있어 독특한 적응력을 나타내고 있다(Fujita et al. 1992; Ichiki et al. 2000; Hwang et al. 2002; Notoya 1978).
이 연구는 무절산호조류 중 동해 연안에 광범위하게 분포하는 수세미말류(Spongites sp.)를 배양하여 포자의 부착과 발아, 생장 경향을 분석하고, 단일 종 배양을 통해 갯녹음 현상의 치유와 복원에 필요한 기술적 이해도를 높이고, 이산화탄소 저감 식물 연구에 활용할 수 있는 기초자료를 제공하고자 하였다.
2. 재료 및 방법
무절산호조류의 채집 및 동정은 강릉시 안현동 해역 조하대에 우점하여 분포하는 무절산호조류가 피복된 암석을 채집하여, 실험실로 이동한 후 생식구조를 분석하였다. 생식기집(conceptacles)의 형태와 영양세포의 구조 관찰을 위해 채집한 무절산호조류의 엽체 표면을 멸균 해수와 브러쉬를 이용하여 수 차례 세척하고, 생식기집이 형성된 부분을 면도칼을 이용하여 기질과 엽체를 분리시켰다. 떼어낸 엽체는 탈회를 위하여 0.6 M nitric acid에 30~60분 동안 침전시켜 준 후, 흐르는 물에 세척하였다. 그 후 5% potassium permanganate (KMnO4)에 20분간 염색을 하고 다시 흐르는 물에서 세척하였다. 탈수를 위하여 30%, 60%, 90% ethanol solution으로 각각 30분간 침전시켜주고 난 후, 100% ethanol solution에 시료를 보관함으로써, cross-section의 전처리 과정을 거쳤다(Harvey et al. 2005). 준비된 시료는 Embedding in Tissue freezing medium으로 고정하여 주고, freezing-microtome을 이용해 10 μm의 두께로 절단하여 광학현미경(Olympus BX50)하에서 관찰하였다.
배양에 사용한 Spongites sp.의 사분포자체는 멸균 해수와 브러쉬를 이용하여 수 차례 세척한 후, 면도칼을 이용해 생식기집이 형성된 부분을 떼어내었고, 떼어낸 모조(about 11 mm2)를 다시 수 차례 세척한 후에 배양에 사용하였다. Tissue culture flask (80×40×20 mm)에 PES배지를 넣어주고, 납작돌잎(Lithophyllum yessoense)의 배양 조건 중 최적 생장 수온인 20℃, 광량 10 μmolㆍm-2ㆍs-1의 조건하에서(Ichiki 2000; Hwang et al. 2002), 30 rpm으로 유지시킨 Shaker위에서 배양을 하였다. 포자의 발아 및 생장은 도립현미경(Olympus IX70)으로 관찰하였고, Digital Microscope Cam으로 촬영하였다. 취득한 영상은 Tomoroscope eye 3.1 소프트웨어를 이용하여 생장률을 측정하였고 3D Surface Plot의 레벨값을 통하여 수직 생장의 여부를 분석하였다.
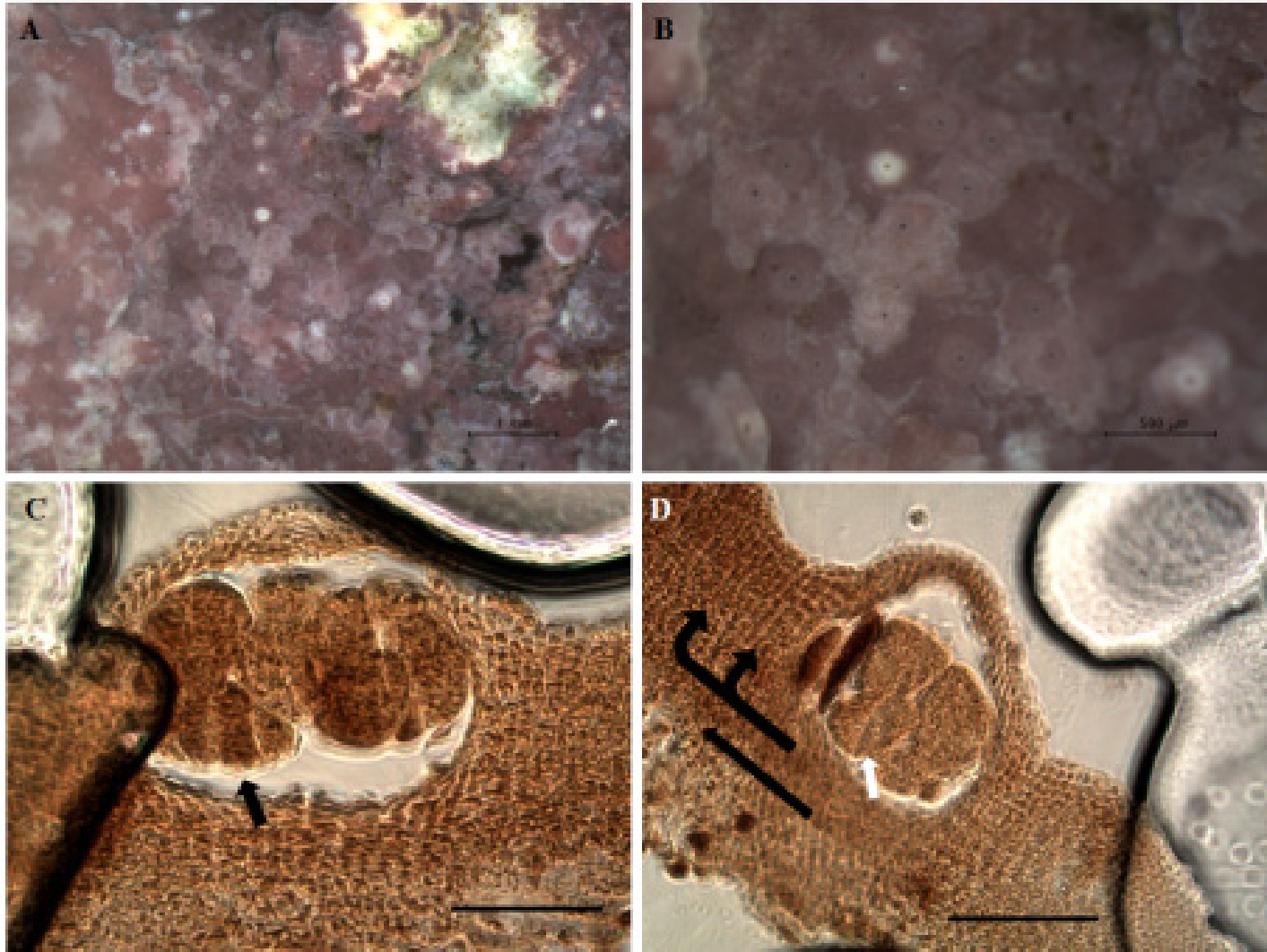
Spongites sp.는 피각화(encrusting)하는 형태로 우점하며 생장하였다. 단공성 생식기집(uniporate conceptacles)을 형성하였고, 생식기집 내부에는 수십여 개의 포자들이 존재하였다. 구형의 포자와 2분열된 이분포자낭(bisporangia) 그리고 띠 모양으로 4분열된 사분포자낭(tetrasporangia)이 존재하였다(Fig. 1). 특히, 단공성 생식기집을 가지며 띠 모양으로 분화된 사분포자낭이 존재하는 것으로써, 채집한 무절산호조류는 Corallinaceae에 속하는 무절산호조류인 것으로 확인되었고, Corallinaceae, Mastophoroideae에 속한 Spongites sp.로 동정되었다.

3. 결과
Spongites sp.는 사분포자체로서 단공성 생식기집을 형성하고 있으며(Fig. 2A, B), 내부구조에서 생식기집은 주변보다 높게 융기되어 형성되었다. 통수관(pore canal)은 아직 형성되지 않은 단계였지만 통수관 주변의 세포가 수평방향으로 형성 되었고, 생식기집 격실(conceptacle chamber)의 가로 직경은 112 μm였고, 격실(chamber) 하부에는 띠 모양으로 분화된 사분포자낭이 55 μm의 길이로 형성되었다(Fig. 2C). 무절산호조류의 몸체를 이루는 엽체의 구조는 수평방향으로 뻗어나가면서 수직방향으로 생장해가는 형태(dimerous thallus)의 패턴으로 나타났고, 표층세포의 형태는 납작하고 둥그런 모양으로 구성되었다(Fig. 2D).
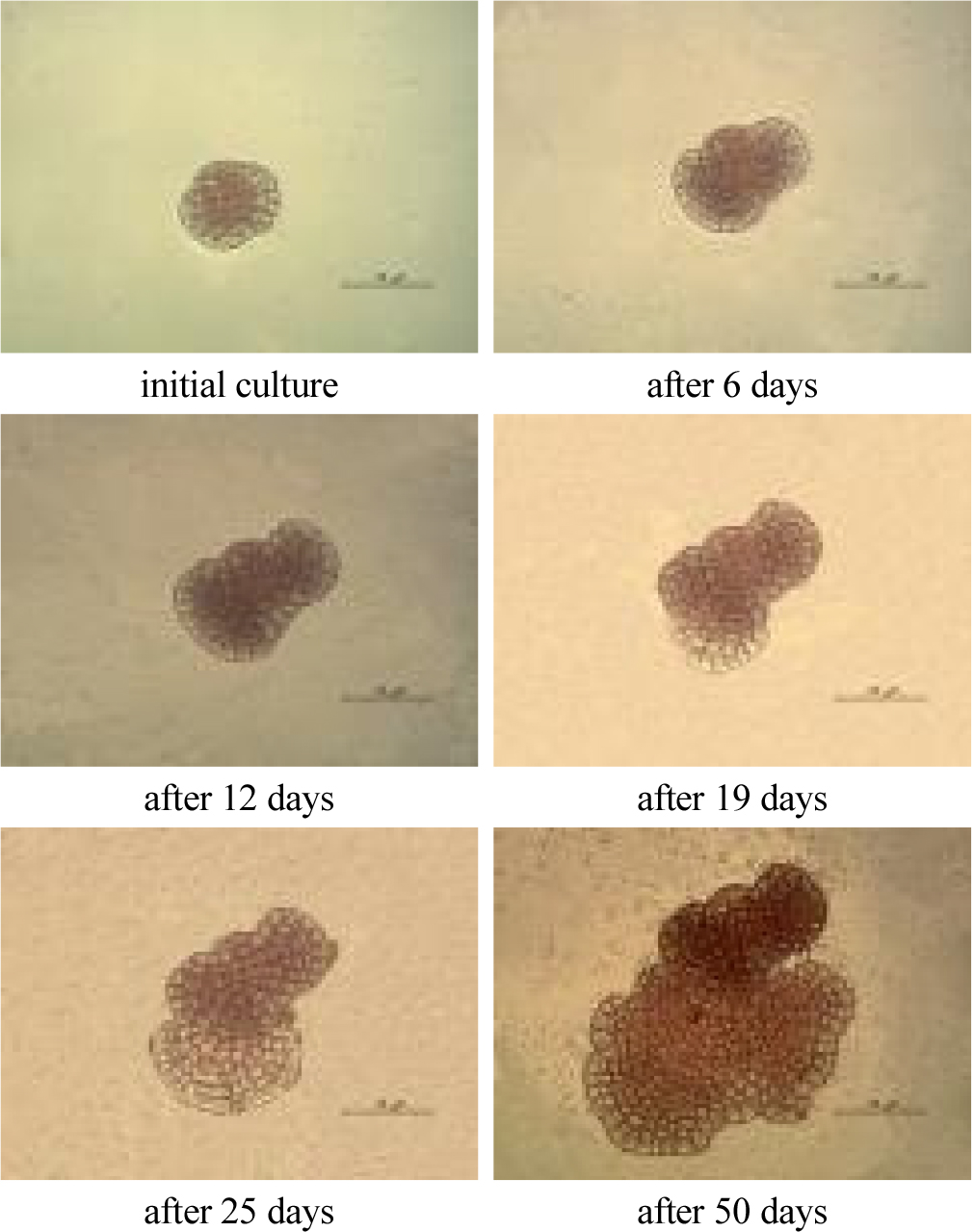
Spongites sp.의 사분포자는 직경 약 30 μm의 구형 형태로 방출되었고, 시간이 경과하면서 포자 주변에 점액질이 포자 주변을 둘러쌓았다. 이는 기질로 착생되는 것을 의미하며 관찰 결과, 점액질이 나타나기 시작하고, 2.5시간부터 4시간 동안에 사분포자 주변을 감싸듯이 형성되었다. 6.5시간이 지나면서 이 물질들은 감소하였고, 26.5시간 이후 완전한 착생이 이루어지면서 대부분 사라지고 점액질은 관찰 되지 않았다.
착생이 완료된 사분포자는 부착한 원세포에서 분화가 먼저 진행되고 그 후에 세포 가장자리의 연변부에서 생장이 진행되는 형태로 발달했다. 1차적으로 가로축과 세로축을 주축으로 십자 형태를 이루어 분화되고, 연변부를 향하여 원호를 그리며 2차, 3차로 분화되는 형태로 발달하였다. 연변세포는 세포내 물질이 비어있는 듯한 형태로 생장이 먼저 진행되고 점차 크기가 증가된 후에 세포내 물질이 이동 또는 생성되는 형태로 발달하였고, 원세포의 색과 동일하였다. 포자는 직경이 45.5 μm이고 엽면적이 1,536.7 μm2이었다. 배양 48시간 후에는 직경 54.9 μm, 엽면적 1,841.9 μm2로 생장하였고, 배양 192시간 후에는 직경이 73.0 μm, 엽면적 2,996.1 μm2로 증가하였다. 여기서 연변부의 세포색이 연한 부분을 제외한 면적은 2,059.8 μm2로써 배양 72시간의 엽면적 1,941.8 μm2, 120시간 후 엽면적 2,387.1 μm2의 중간값으로 세포질 내 물질들이 점차적으로 이동 또는 생성되었다.
발아한 세포에서 배양일 수에 따라 각각의 생장 단계를 살펴보면, 발아 단계에서와 비슷한 유형의 한 방향성 또는 두 방향성으로 생장하였다. 실험실 배양 조건과 환경에 따라 생장 유형이 다양하게 변화하지만, 본 배양 조건에서는 위와 같은 두 가지 형태의 유형으로 생장하였다(Figs 3, 4).

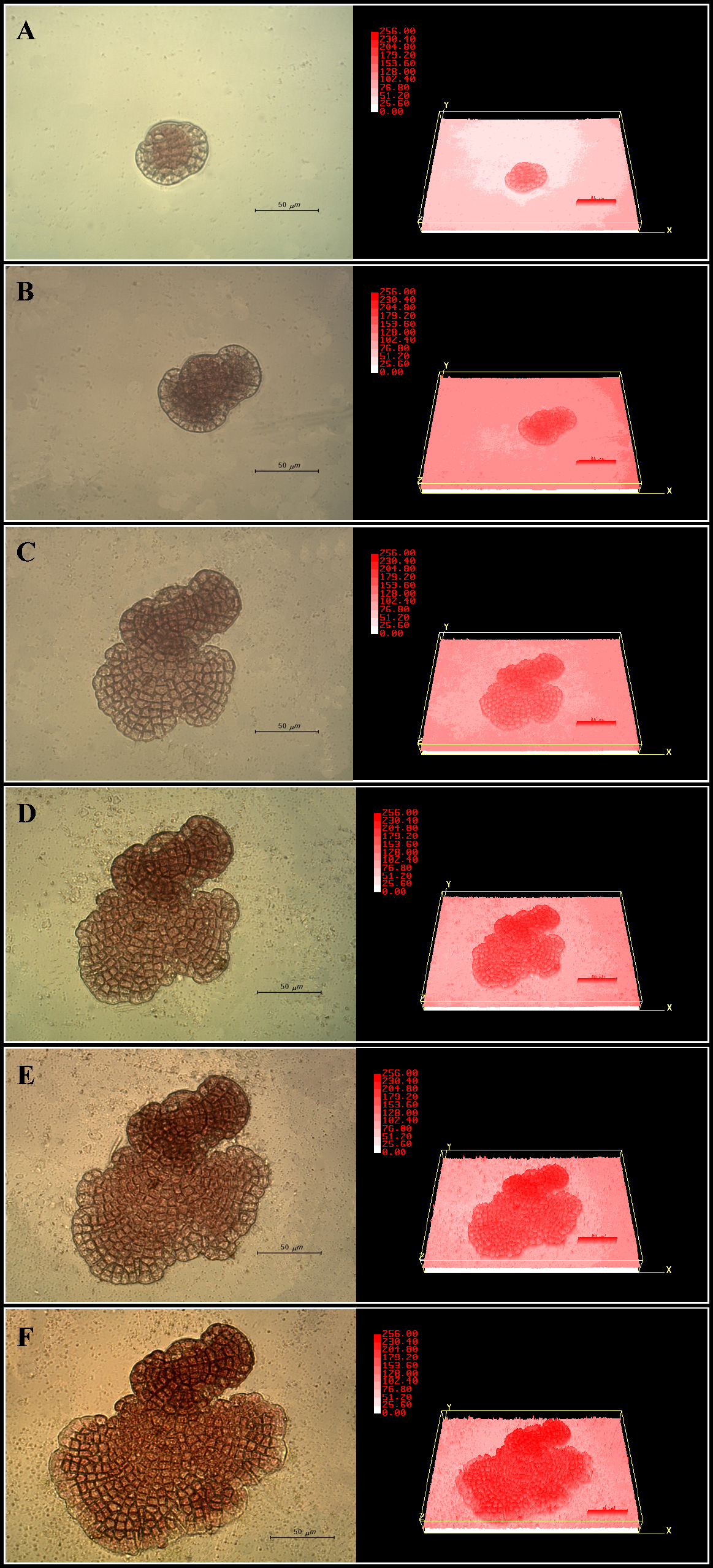
Spongites sp. 사분포자들은 부착하여 분화가 진행되었고, 분화하는 모든 세포들은 중앙의 원세포를 중심으로 각각의 방향성을 가지고 주변으로 확장해 나가며 생장이 진행되었고 엽면적(μm2)의 증가량은 Fig. 5와 같다. 일일 생장율(Fig. 6), 배양 후 18일까지 일일 생장율에서 수치상 변화폭은 나타나지만, 유의적인 차이를 보이지 않았다. 19일차에 접어들면서 생장율이 크게 증가하였고, 배양 21일 이후부터 43일까지 일일 생장율이 약 500 μm2 day-1 수준으로 유지되다가 50일경 다시 생장율이 증가한 후 다시 감소하는 경향을 나타냈다. 피도 레벨을 이용하여 3차원 이미지로 구현한 3D surface plot level (SPL)으로 수직 생장이 이루어진 것을 확인할 수 있었다(Fig. 7). 배양 초기부터 36일까지 엽면적은 증가되었으나, SPL값은 큰 변화를 나타내지 않았다. 배양 43일째에 접어들면서 엽면적의 소폭 증가와 SPL값 증가(179.20 ~ 204.80)를 통해 수직 생장이 이루어졌다는 것을 확인하였다. 배양 50일차에는 엽면적 증가와 함께 초기에 생성된 세포의 부위에서만 SPL값이 204.80 ~ 230.40으로 증가하였다. 배양 57일째로 접어들면서 4,911.6 μm2의 엽면적 증가가 이루어졌고, 원세포와 연변부의 세포 모두에서 SPL값이 230.40으로 증가되어 수평과 수직 생장이 모두 진행된 것을 확인하였다.


4. 고찰
무절산호조류(crustose coralline algae)는 엽상 해조와 함께 연안 생태계에서 광합성을 하는 1차 생산자이며, 세포벽에 탄산염을 다양한 형태로 축적시키는 해양생태계에서 중요한 식물로 알려져 있다(Johansen 1981). 자연 환경에서 무절산호조류는 엽상 해조의 부착기 때문에 쉽게 눈에 띄지 않지만, 엽상 해조가 유실되면 최하부에 피복하며 생장하던 무절산호조류가 드러나 쉽게 관찰된다. 생태적 서식 특징으로 무절산호조류의 피복도를 기준으로 갯녹음 현상의 지표종으로 활용되고 있다(Park 1976; Hwang et al. 2002; MOF 2002; Kim 2006; NFRDI 2008; Park 2008; Chung et al. 1998). 무절산호조류의 표층세포에서 발현되는 탈각작용(Keats and Maneveldt 1994; Keats et al. 1997) 및 납작혹돌잎속(Lithophyllum spp.) 표층에 착생한 다시마(Saccharina Japonica) 유주자 발아가 저해(Masaki et al. 1981)된다는 연구 등에 의해 무절산호조류를 바라보는 부정적 인식이 나타나기도 한다. 하지만, 자연적인 미소서식처에서 무절산호조류가 생장하고 있는 표층에 다시마(S. japonica)와 미역(Undaria pinnatifida), 괭생이모자반(Sargassum horneri) 등 대형 갈조류 포자체(sporophyte)들이 서로 공존하며 생장하기도 한다. 탈각 및 저해 현상이 무절산호조류 주변에서 나타나기는 하지만 무조건 엽상 해조의 생장제한 관련된 원인을 제공하지는 않는 것으로 보이며, 무절산호조류에 따라 차이가 있으며, 생장과 생식에 장해가 될 때 아직 명확하지 않은 기작이 일시적으로 발현되는 것으로 판단된다.
동해 연안 조하대 지역에 광범위하게 분포하는 무절산호조류 중 Spongites sp.의 사분포자는 방출 후 약 24시간이 되면, 기질에 완전한 착생이 이루어지고, 세 가지 형태로 분화가 이루어졌다. 무절산호조류의 생활사는 홍조류에서 보편적으로 나타나는 무성세대의 사분포자체와 유성세대의 암ㆍ수 배우체가 동형의 형태로 교번하는 것으로 알려져(Johansen 1981; Harvey et al. 2005), 다른 형태로 분화되는 패턴은 암ㆍ수 배우체와 무성의 단포자 등이 다른 형태로 분화가 이루어지는 가능성도 생각해 볼 수 있다. 포자의 분화 속도는 전반적으로 느리게 진행되었고, 발아하지 않고 분화가 먼저 진행되는 형태의 포자에서 생장이 가장 빠르게 진행되었다. 이러한 무절산호조류의 포자발아 패턴은 Notoya (1974)와 Chihara (1974)에 의해 보고되었는데, 본 연구에서도 Spongites sp.의 포자발아 패턴이 앞서 연구된 결과와 유사하게 관찰되었고, 그 중 Chihara (1974)가 제안한 Fosliella farinosa-type과 가장 유사한 형태로 분열되었다. 생장 경향은 계단식 구조로 진행이 되는 것으로 추정되었다. 이는 기본적으로 수평과 수직 생장이 함께 진행될 때는 생장율이 낮고, 수직 생장이 둔화되었을 때에 생장율이 급격히 증가되는 결과에서 유추할 수 있으며, 무절산호조류 영양세포의 구조적 특징인 수평 생장과 수직 생장의 형태(nomerous or dimerous)를 Spongites sp.의 배양연구를 통해 입증할 수 있었다.
본 무절산호조류의 배양 연구를 통하여 갯녹음이라는 전 세계적인 생태계 이상 현상을 이해하는 기초 자료의 구축에 기여하였다고 판단되며, 향후 이산화탄소 흡수력, 엽상 해조와의 상호관계 파악 등의 연구에 활용 가능성 높은 자료가 될 것으로 판단된다.
