

1. 서론
2. 재료 및 방법
2.1 해조류 이식로프 제작 및 인공기질 이식
2.2 연구 정점
2.3 이식해조류 모니터링 및 분석
3. 결과
3.1 이식 해조류의 생존률
3.2 이식 해조류의 생장
4. 고찰
1. 서론
해조류 군락은 해양생태계의 가장 기초적인 생산자 역할 뿐만 아니라 주변의 다양한 생물에게 에너지원, 산란장, 은신처 및 생육장소를 제공하여 해양의 생물다양성 유지에 기여한다(Steneck et al. 2002). 또한, 일부 해조류 종들은 오래 전부터 인간사회에 식품, 약재 및 비료 등 유용한 자원으로 활용되어 왔으며, 이들이 함유하고 있는 각종 유용 물질을 기반으로 의약, 사료, 향장 및 생물연료개발 등 고부가가치 창출을 위한 다양한 연구 및 산업활동도 증가하고 있다(Garcia-Vaquero and Hayes 2016; Kinley et al. 2016; Ryu et al. 2018; Suutari et al. 2015; Yun et al. 2015). 특히, 해중림의 주요 구성원인 대형갈조류 군락은 막대한 생물량을 바탕으로 대기 중에서 해양으로 유입된 이산화탄소를 효과적으로 고정할 수 있다. 최근의 연구에 의하면, 전 세계 연안의 모자반류(Sargassaceae)가 고정하는 탄소량은 맹그로브(Mangrove), 염생식물(Salt marsh) 및 해초류(Seagrasses)의 탄소고정량을 합한 것을 능가할 것으로 추정된다(Gouvêa et al. 2020).
제주도 조하대 천연 해조류 군락의 층상구조는 주로 다년생 모자반류가 최상층에서 수관(Canopy), 그 하부에 다년생 대형갈조류인 감태(Ecklonia cava)가 준수관(Sub-canopy), 저층에는 대마디말류(Cladophora spp.), 우뭇가사리류(Gelidium spp.), 곱슬이류(Plocamium spp.) 등 다양한 중소형 해조류와 함께 유절산호말류(Articulated coralline algae)가 군집을 이룬다(Kang et al. 2011; Kang and Kim 2012). 특히, 감태와 모자반류 같은 대형 갈조류들의 넓은 체표면적과 복잡한 구조는 새붉은실류(Melanothamnus spp.), 민털이풀류(Anotrichium spp.) 및 비단풀류(Ceramium spp.) 등 소형 해조류들이 착생할 수 있는 기질을 제공하고, 빛에 민감한 해조류 종들의 그늘막 역할을 하여 해조류의 종다양성 유지에 기여한다(Kang and Kim 2013; Kang et al. 2019). 또한, 대형갈조류는 과거로부터 오랫동안 제주도 주민의 귀중한 식량 및 비료자원으로 활용되어 왔으며, 현재에도 모자반(Sargassum fulvellum), 경단구슬모자반(S. muticum), 톳(S. fusiforme) 및 감태(E. cava) 등의 갈조류는 홍조류인 우뭇가사리(Gelidium elegans)와 함께 제주도 어촌 구성원들의 중요한 경제적 자원으로 이용되고 있다(Kang et al. 2019).
해조류가 해양 생태계 및 인간사회에 미치는 중요한 역할에도 불구하고, 전 세계에 보고되고 있는 해조류 군락의 급격한 감소 현상은 제주도 연안을 포함한 우리나라 전 연안에서도 광범위하게 발생하고 있다. 이 현상은 다년생 대형갈조류 군락으로 대표되는 해중림(Marine forest)의 소실을 동반하며, 심할 경우 저층의 중소형 해조류는 물론 유절산호말류 군락까지 탈락하여 최하층의 무절산호말류(Melobesioidean algae)가 하얗게 드러나는 이른바 갯녹음 현상(Sea barren or isoyake)으로 나타난다(Campbell et al. 2014; Chung et al. 1998; Fujita 2010; Hwang et al. 2017; Jung et al. 2019). 이러한 해조류 군락의 감소현상을 유발하는 원인으로는 지구온난화에 의한 해수온 상승(Hwang et al. 2017; Serisawa et al. 2004), 조식동물의 먹이활동 증가(Jeon et al. 2015; Nimura et al. 2007), 해안 개발에 의한 부유물의 유입(Fujita 2010; Kim et al. 1996), 도시화 진행에 따른 오염물질 유입의 증가(García et al. 2020; Scherner et al. 2013) 등 다양한 요인들이 거론되고 있다.
해조류 군락 소실 현상의 대응책으로 우리나라에서는 해조류 군락 동태에 대한 지속적인 모니터링(Jung et al. 2019; Kim et al. 2013a), 갯닦기, 투석 및 해중림초 설치 등 기질전환(Heo et al. 2015; Kim et al. 2013b; Kwak et al. 2014), 조식동물 제거(Jeon et al. 2015), 포자주머니 설치(Choi et al. 2000) 등 다양한 방법을 시도하고 있다. 또한, 수중 저연승 시설에 해조류를 부착 및 생육시켜 포자확산을 유도하거나 배양된 대형갈조의 배아 또는 유엽을 인공기질에 부착시켜 해중림을 조성하는 등 해조류를 직접 이식하는 방법도 제시되고 있다(Choi et al. 2002, 2003; MIFAFF and NFRDI 2010; Yoon et al. 2014). 그러나 발아 초기 유엽상태인 해조류는 조식동물의 좋은 먹이표적이 되므로 이들의 식해에 대한 대책으로서 Choi et al. (2002)은 성체 해조류 이식을 제시하기도 하였다. 성체 해조류를 인공기질에 이식하는 방법으로는 수지상 부착기를 케이블타이(Zip-tie)로 로프에 고정하여 이식 로프를 만든 후 인공기질에 고정하거나 인공기질에 미리 로프를 부착한 후 해조류를 수중에서 케이블타이로 고정하는 방법이 주로 활용되고 있다(FIRA 2019). 그러나 케이블타이 고정방법은 커다란 수지상 부착기(finger- like holdfast)를 가진 감태, 곰피(Ecklonia stolonifera), 대황(Eisenia bicyclis), 다시마(Saccharina japonica) 등의 해조류 종에 한하여 사용할 수 있지만, 반상(Discoid) 또는 원추상(Conical) 부착기(holdfast)를 가진 대부분의 모자반류에는 적용하기 어렵다.
이 연구는 인공기질에 원추상 부착기를 가진 다년생 모자반류 성체를 고정하여 이들의 생존률과 생장양상을 모니터링 하고, 해조류 성체 이식에 주로 사용되고 있는 케이블타이 고정 방식과 해조류 부착 효과성을 비교하여, 이 연구에서 사용한 다년생 모자반류 성체 이식 방법의 해중림 조성 적용 가능성을 탐색하고자 수행되었다. 또한, 감태 또는 다년생 모자반류 단일 분류군을 이식한 그룹과 두 분류군을 혼합하여 이식한 그룹 간의 이식효과도 비교하였으며, 이식된 해조류의 부착 안정성을 시험하기 위하여 물리적 교란을 반영할 수 있는 장소(수심 5 m 내외, 태풍 영향 등)에 설치하였다.
2. 재료 및 방법
2.1 해조류 이식로프 제작 및 인공기질 이식
이식에 사용한 감태와 큰열매모자반은 2018년 8월 서귀포시 신양리 조하대 수심 10~15 m에서 엽장의 길이가 20~30 cm인 개체들을 채집하였다. 채집 시 끌칼 등으로 부착기가 손상되지 않도록 유의하였으며, 채집된 개체들은 해수에 보관한 후 이튿날 해조류 이식로프 제작에 사용하였다. 해조류를 부착시키기 위한 로프는 지름이 20 mm인 PE (Polyethylene) 재질이며, 1 m씩 절단하여 사용하였다.
감태를 고정하기 위한 로프에는 10 cm 간격으로 총 10개의 이식 위치를 페인트로 표시하였다(Fig. 1A). 큰열매모자반을 고정하기 위한 로프는 해조류 이식 하루 전 약 3 x 3 cm2 크기의 단추 형태로 성형된 에폭시 수지(E-380, Konishi, Japan)를 로프 위에 10 cm 간격으로 10개씩 부착한 후 경화시켰다(Fig. 1B). 이 연구에서 사용된 에폭시 수지는 반고체상의 흰색 주제와 황록색 경화제로 구성되어 있고, 이 두 용제를 잘 섞으면 흰색의 반고체 접착제로 변한 뒤, 24시간 후 완전히 경화되어 딱딱한 고체가 되는 특성이 있다. 감태 및 큰열매모자반 혼합 이식을 위한 로프는 10 cm 간격을 두고 교대로 감태 부착 지점 표시 및 단추 형태의 에폭시 수지를 부착하였다(Fig. 1C).

Fig. 1
The procedures for transplantation of Ecklonia cava and Ecklonia cava on the artificial reef in this study. (A) Mark (red arrowheads) at 10 cm interval on PE lope for the position to attach E. cava. (B) Attached harden epoxy resin (red arrows) at 10 cm interval on PE lope for attaching S. macrocarpum. (C) Epoxy resin (arrow) and mark (arrowhead) on rope for combining E. cava and S. macrocarpum. (D) Smoothing S. macrocarpum holdfast with knife. (E) Air drying S. macrocarpum holdfast. (F - G) Attaching S. macrocarpum holdfast on the harden epoxy resin with instant glue. (H) Fixation of E. cava with zip tie. (I) Completed transplantation rope for E. cava group (upper), combined group (middle), and S. macrocarpum group (lower). (J) Fixed transplantation rope on the artificial reef.
이식할 큰열매모자반은 접착부위가 편평하도록 부착기 바닥면을 면도날로 얇게 잘라내어 이물질 등을 제거하였고, 에어건으로 접착면의 습기를 제거한 후 순간접착제를 도포하여 이식로프 상에 부착되어 있는 에폭시 수지에 고정하였다(Fig. 1D-G). 감태는 수지상 부착기를 로프에 표시된 부착부위에 올려놓고 케이블타이로 고정하였다(Fig. 1H). 감태 단일 분류군 이식로프, 큰열매모자반 단일 분류군 이식로프 및 감태/큰열매모자반 혼합 이식로프를 각각 4개씩 제작하였으며, 각 이식로프에는 10 개체씩 대상 해조류가 부착되었다(Fig. 1I). 따라서 이 연구에서 이식된 해조류는 감태 60개체(단독 이식군 40개체, 혼합 이식군 20개체) 및 큰열매모자반 60개체(단독 이식군 40개체, 혼합 이식군 20개체) 등 총 120개체이다.
해조류를 이식할 인공기질은 동일 평면상에 3개의 돌출부가 있는 콘크리트 재질의 트라이포드(Tripod)로서 하나의 돌출부는 길이 약 1 m, 폭 약 50 cm로 가장자리를 따라 비스듬한 경사면이 형성된 형태이며 저층으로부터 상단면까지 높이는 약 50 cm이다. 해조류 이식로프의 인공기질 부착은 수중에서 공기통이 결합된 타정기(Air nailer)를 이용하였다. 인공기질 1기의 돌출부 3개에 각각 감태 단독 이식군(Ecklonia group), 큰열매모자반 단독 이식군(Sargassum group) 및 감태/큰열매모자반 혼합이식군 (Combined group) 1개씩, 총 4기의 인공기질에 부착하였다(Fig. 1J).
2.2 연구 정점
해조류 이식 실험은 제주도 동쪽에 위치한 종달항 인근으로, 해안선으로부터 약 300 m 외해에 위치하여 있으며 수심은 최간조 시 약 5 m 내외이다(Fig. 2). 저층은 현무암으로 구성된 평암반 지대로 곳곳에 모래가 얕게 퇴적된 지형이 산재하고 있다. 해조류 군락은 감태가 최우점 종으로 준수관을 이루고 있고, 유절산호말류(Articulated coralline algae)가 준우점하면서, 갈래곰보(Meristotheca papulose), 두가닥바닷말류(Dichotomaria spp), 볏붉은잎(Callophyllis japonica), 붉은뼈까막살(Grateloupia angusta) 및 우뭇가사리(Gelidium elegans) 등의 중형 해조류가 하층에 산재하여 분포하고 있다. 이 지역에서 관찰된 조식동물은 주로 소라(Turbo cornutus, Turban shell)와 보라성게(Heliocidaris crassipina, Sea urchin) 두 종류였으며, 모두 1 ind./m2 미만으로 드물게 출현하였다. 연구정점의 반경 50 m 이내에 감태를 제외한 모자반류 등 다른 대형갈조류의 서식은 관찰되지 않았다.
2.3 이식해조류 모니터링 및 분석
인공기질 위 해조류 이식 로프 부착은 2018년 8월에 수행하였으며, 부착작업을 하는 동안 큰열매모자반 단독 이식군에서 2개체가 탈락하였고, 감태/큰열매모자반 혼합 이식군에서 감태 1개체가 탈락하였다. 따라서, 생존률 및 생장 모니터링은 감태 단독 이식군 40개체, 큰열매모자반 단독 이식군 38개체 및 혼합 이식군 39개체(감태 19, 큰열매모자반 20)를 대상으로 수행하였다.
해조류 이식 후, 이식된 개체마다 고유 식별번호를 부여하여 수중에서 엽체의 길이 및 탈락상태를 기록하였다. 모니터링은 매월 반복 측정을 기본으로 하였으며, 태풍의 직접적 영향 직후에도 추가적으로 수행하였다. 해조류 생존률은 인공기질에 부착된 상태를 유지한 개체 수를 근거로 산출하였다. 산출된 생존률 값을 통해 두 가지 이식방법 즉, 케이블타이 고정법을 이용한 감태이식 및 에폭시 수지와 순간접착제를 이용한 다년생모자반류 고정법 간의 생존률을 비교하였다. 또한, 3가지 이식군(감태 단독, 큰열매모자반 단독 및 감태/큰열매모자반 혼합) 사이의 생존률도 비교하였다. 엽체 길이는 감태의 경우 부착기 상부부터 중심 가지의 정단부까지, 큰열매모자반의 경우 부착 기질부터 엽체의 정단부까지의 길이를 1 cm 단위까지 측정하였다(Choi et al. 2015). 해조류 생장 양상은 시험이 종료될 때까지 인공기질에 부착상태를 유지하고 있는 이식 해조류들 중 각 분류군에서 가장 크게 자란 5개체를 선별하여 분석하였다. 이식 해조류의 개체 수 및 엽체 길이를 측정하는 동안 이식된 해조류의 생식기관 형성 유무 및 부착기 발달 양상을 함께 관찰하여 기록하였다.
3. 결과
3.1 이식 해조류의 생존률
2018년 8월 10일 최초 이식 후 제주도 연안은 8월 23일과 10월 6일에 최대풍속 30 m/s를 초과하는 강한 바람을 동반한 태풍 제19호 솔릭(Soulik)과 제25호 콩레이(Kong-rey)의 직접적인 영향을 받았다. 이 외에도 8월 15일 제15호 리피(Leepi), 9월 4일 제21호 제비(Jebi) 및 9월 30일 제24호 짜미(Trami) 등 강한 태풍의 간접 영향을 받아 해조류 이식 장소에 대한 크고 작은 물리적 교란이 수 회 발생하였다(KMA, 2018).
1차 모니터링은 제15호 태풍 리피의 간접 영향과 제19호 솔릭의 직접 영향 직후인 8월 25일에 실행하였으며, 그 결과 감태 단독 이식군 19개체, 큰열매모자반 단독 이식군 15개체 및 혼합 이식군 9개체(감태 7, 큰열매모자반 2)가 탈락한 것으로 관찰되었다. 2차 모니터링은 제21호 태풍 제비의 간접 영향 후인 9월 15일에 실행하였다. 그 결과 감태는 단독 이식군에서 1개체 및 혼합 이식군에서 2개체가 추가로 탈락하였으며, 큰열매모자반은 단독 이식군에서 2개체가 탈락하였다. 3차 모니터링은 제24호 태풍 짜미의 간접영향과 제25호 콩레이의 직접영향 후인 10월 8일에 실행하였으며, 그 결과 감태 단독 이식군에서 1개체, 큰열매모자반 단독 이식군에서 5개체, 혼합 이식군에서 1개체(감태 0, 큰열매모자반 1)가 추가로 탈락하였다. 그 후 실험종료 시까지 큰열매모자반은 단독 이식군 및 혼합이식군에서 추가로 탈락한 개체는 관찰되지 않았으나, 감태의 경우 단독 이식군에서 추가적인 탈락이 지속적으로 발행하여 최종 11 개체가 관찰되었다.
케이블타이 고정법을 이용하여 이식한 감태 59 (단독 이식군 40, 혼합 이식군 19)개체와 에폭시 수지와 순간접착제를 이용하여 이식한 큰열매모자반 58 (단독 이식군 38, 혼합이식군 20)개체의 생존률을 비교하였을 때, 태풍에 의한 물리적 교란이 발생하는 동안 두 분류군 모두에서 급격한 생존률 감소가 나타났다. 그러나 마지막 태풍의 영향 후, 이식된 감태의 개체 수는 지속적으로 감소하는 반면, 큰열매모자반의 개체 수는 실험종료 시까지 지속적으로 유지되어 1년 후 큰열매모자반 56.9%, 감태 28.8%로 큰열매모자반을 에폭시 수지와 순간접착제를 이용한 이식 방법에서 더 높은 생존률을 나타내었다(Fig. 3A).
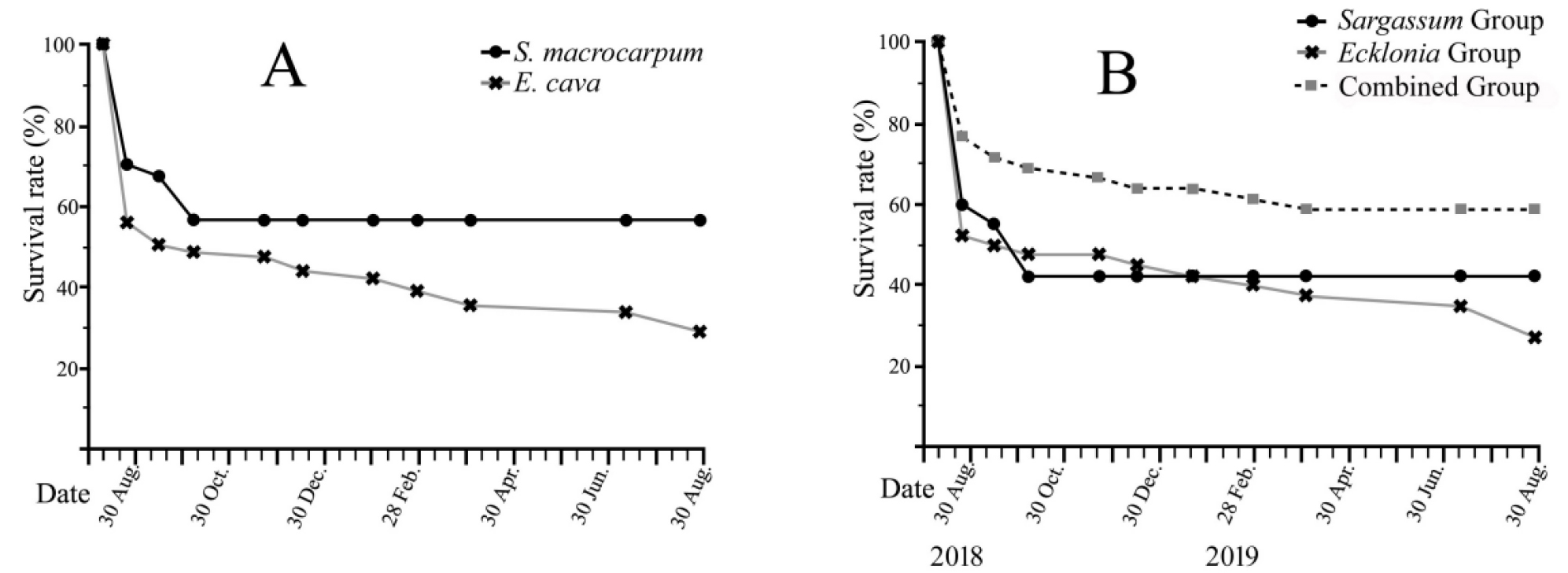
Fig. 3
Survival rates of transplanted Ecklonia cava and Sargassum macrocarpum. (A) Comparison of E. cava (black crosses on gray line, n = 59: 40 Ecklonia-group and 19 combined group) and S. macrocarpum (black circles on black line, n = 58: 38 Sargassum-group and 40 combined group). (B) Comparison among the groups for Ecklonia (black crosses on gray line, n = 40), Sargassum (black circles on black line, n = 38), and combined (gray squares on gray spot line, n = 39: 19 Eklonia cava and 20 Sargassum macrocarpum).
감태 단독 이식군, 큰열매모자반 단독 이식군 및 감태/큰열매모자반 혼합 이식군 등 3가지의 이식군을 비교한 결과 감태 및 큰열매모자반 단독 이식군은 태풍에 의해 대량 탈락하여 10월까지 50% 이하의 생존률을 보인 반면, 혼합 이식군은 66.7%로 2개의 단독 이식군보다 더 높은 생존률을 보였다. 최종 생존률은 감태/큰열매모자반 혼합 이식군에서 59.0%로 가장 높게 나타났으며, 큰열매모자반 단독 이식군 42.1%, 감태 단독 이식군 27.5% 순으로 나타났다(Fig. 3B, Fig. 4).

3.2 이식 해조류의 생장
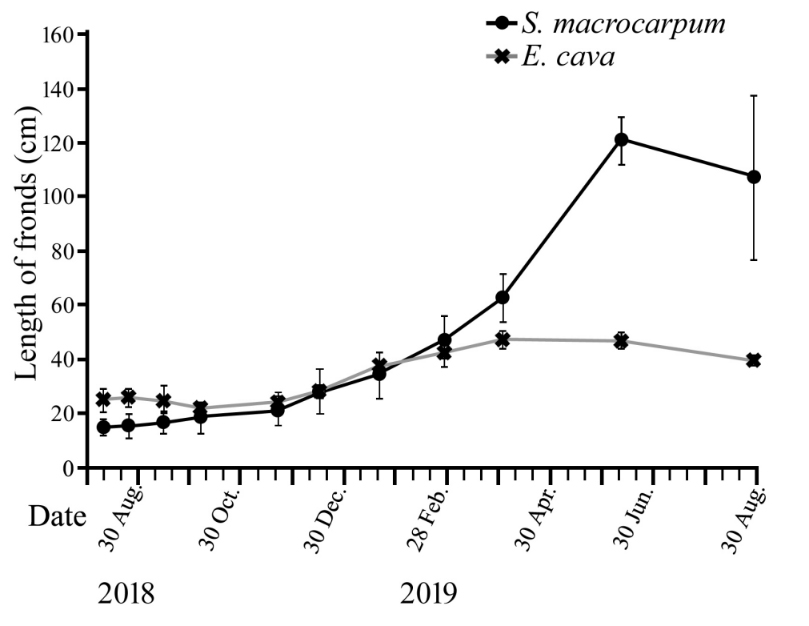
인공기질에 이식된 감태의 엽체 길이는 이식 후 3개월까지 큰 변화 없이 유지되다가 2018년 12월부터 완만한 증가를 보이며, 초기 평균 24.9 cm가 2019년 4월에 평균 47.5 cm까지 생장한 후 6월까지 그 크기가 유지되었다. 이식 12개월 후인 2019년 8월에는 6월 보다 다소 감소한 평균 39.7 cm로 측정되었다. 큰열매모자반의 엽체 길이는 이식 3개월 후인 2018년 11월까지 다소 완만한 생장을 한 이후 점점 빠른 생장을 보였고 2019년 4~6월에 급격히 생장하여 약 2개월만에 엽체 길이가 평균 62.7 cm에서 120.9 cm로 거의 두 배가 되었다. 그 후 엽체 상부가 노성해지면서 8월에는 길이가 다소 감소하였다(Fig. 5).
이식된 감태의 부착기는 이식 1개월 후 기존 부착기의 말단이 점점 확장하며 이식 4개월 후 로프를 넘어서 인공암반에 부착하기 시작하였으며(Fig. 6A-B), 그 후 새로운 부착기가 줄기의 기부에서 형성되기 시작하여 이식 8개월 후 기존 부착기를 완전히 덮으며 말단부가 인공기질에 완전히 부착하였다(Fig. 6C). 큰열매모자반의 부착기는 이식 1개월 후 부착면의 절단 부위에 상처가 아무는 형태로 새로운 조직들이 자라는 것이 관찰되었으며, 이식 4개월 후 절단면의 상처가 완전히 아물며 단추형 에폭시 수지 위에 단단히 고정되었다(Fig. 6D-E). 이식 8개월 후 부착기는 단추형 에폭시 수지의 상단면을 거의 완전히 덮어 이식로프까지 확장되었다(Fig. 6F).
큰열매모자반의 어린 생식기탁은 6월에 발생하여 8월에 완전히 성숙하였으며(Fig. 7A-B), 감태는 8월에 성숙한 포자낭반을 관찰할 수 있었다(Fig. 7C). 이듬해 6월 인공기질 위에 새로 착생한 수 개체의 감태(Fig. 7D)와 큰열매모자반(Fig. 7E) 어린 개체들이 관찰되었다.


Fig. 7
Reproduction of transplanted Sargassum macrocarpum (A, B, E) and Ecklonia cava (C, D). (A) Young conceptacles (arrows) on the branchlets of S. macrocarpum on 11 Jun. 2019. (B) Mature male conceptacles (arrows) of S. macrocarpum on 23 Aug. (C) Sporangial sori on the blade of E. cava on 23 Aug. (D-E) Young frond of E. cava (D) and S. macrocarpum (E) on artificial reefs on 22 Jun. 2020.
4. 고찰
연안의 대형갈조류 숲을 포함한 해조류 군락의 쇠퇴에 대응하기 위해 우리나라에서는 과거 수십 년 전부터 바다숲 조성사업을 지속적으로 시행하여 왔으며, 인공기질과 해조류 이식을 결합한 방법은 주요 이식 대상종으로서 감태, 곰피, 대황, 다시마 등 엽상형 다년생 대형갈조류와 모자반(Sargassum fulvellum)이 선정되어 왔다(FIRA 2015; Kim et al. 2007, 2012; MOF and FIRA 2015). 이들 종 대부분이 경제적인 유용성으로 인해 오래 전부터 대량 양식 기술이 정립되어 있고, 커다란 수지상 부착기를 갖는 형태적 특징을 지니고 있어서 로프 위에 철사나 끈 또는 케이블타이로 고정하기가 용이하기 때문으로 여겨진다(Hwang and Park 2020; Kang 1968; Kim et al. 2017).
한정된 해중림 조성 대상종의 다양성을 높이고자 괭생이모자반(Sargassum horneri), 큰열매모자반(S. macrocarpum), 외톨개모자반(Myagropsis myagroides) 등 대형갈조류 종들을 해중림 조성에 적용하려는 연구가 최근 수행되고 있다(Choi et al. 2003; FIRA 2017; Ko et al. 2020). 그러나 괭생이모자반은 수심에 따라 높이 10 m까지 자라 무성한 모자반 숲을 조성할 수 있다는 장점이 있는 반면, 단년생(annual)이기 때문에 인공기질에 이식한 후 초여름에는 기질에서 탈락하여 조성된 해중림을 꾸준하게 유지하지 못하는 단점이 있다(Kang et al. 2019). 특히, 최근 중국에서 대량 발생하여 우리나라 연안에 갈조류 대발생(Golden-tide)을 유발함으로써 선박안전을 저해하고 천해양식 시설에 피해를 줌과 동시에 대량의 해안 쓰레기를 유발하는 등 부정적인 사회적 인식이 있어 해중림 조성에 적용하기 곤란한 점이 있다(Byeon et al. 2019; Hwang et al. 2016). 한편, 인공배양을 통한 큰열매모자반 또는 외톨개모자반을 이식하는 방법은 현재 초기 연구단계로서 인공증식 체계가 확실하게 정립되어있지 않은 상태이다(FIRA 2017; Ko et al. 2020). 앞서 거론되었던 조식동물의 좋은 먹이 표적이 되는(Choi et al. 2002) 어린 개체군들을 보호할 방안도 마련되어야 한다(Yoon et al. 2014). 다년생 모자반류의 포자 방출시기를 고려하여 로프를 포함한 구조물을 모자반류 군락 인근 저층에 설치함으로써 유배 자연 채묘 및 양성 방안도 제시되었으나(FIRA 2017), 파도 등 수중의 물리적 환경에 의한 유배 채묘 시설의 안정성을 유지하기 어려운 단점이 있다.
우리나라에서 대형 갈조류 성체를 순간접착제를 이용하여 이식하는 연구는 앞서 Choi et al. (2002)에 의해 처음 시도되었다. 그러나 이들이 사용한 대상종은 곰피로서 커다란 수지상 부착기와 기는줄기(Stolon)를 갖는 특성으로 인해 기질에 부착할 수 있는 접착면이 넓기 때문에 부착이 용이하다. 그러나 이 연구에서 사용한 큰열매모자반의 경우 부착기 직경이 2 cm내외로 기질 접착면이 매우 협소한 특징을 갖고 있다. 이 실험에 앞서 실험 전 탐색단계로서 큰열매모자반의 부착기를 PE 재질의 로프 홈에 순간접착제로 직접 부착하는 방법을 사용하였으나 수중에서 운반하는 동안 엽체가 로프에서 쉽게 탈락하는 점을 발견하였다(JGEC 2017). 이에 대한 보완으로 이번 연구에서는 에폭시 수지를 성형하여 PE 로프에 부착시킨 후 경화시켜 큰열매모자반의 부착기를 순간접착제로 에폭시 수지 위에 부착시킴으로써 수중 운반 시 엽체가 탈락하는 문제점을 해결할 수 있었다.
이 연구에서 이식된 큰열매모자반의 가을부터 초겨울까지의 생장 지체 기간을 보낸 후 봄부터 초여름까지 급격히 생장하여 생식기탁을 형성하는 패턴은 자연 군락지에서 생육하는 개체군들과 유사함을 보여주었다(Fig. 2-7, Murase and Kito 1998). 또한, 수심 5 m 이내 비교적 얕은 수심에서 파도 등 높은 물리적 교란과 더불어 이식 초기에 겪은 직간접적인 태풍의 영향에도 기질에서 탈락하지 않고 이식 후 1년 이상 정상적으로 생존한 개체가 57%에 달하는 것으로 나타났다(Fig. 3A). 이러한 결과는 에폭시 수지와 순간접착제를 이용한 이식 방법의 부착강도가 원추형 가근을 가진 대형갈조류 이식에 유효하다는 것을 시사하고 있다. 또한, 여름부터 초가을까지 태풍 영향이 빈번한 시기를 피하여 이식한다면 더 높은 생존률을 나타낼 것으로 판단된다. 단일 분류군으로 구성된 실험군에서, 큰열매모자반 단독 이식군과 감태 단독 이식군을 비교하였을 때, 태풍의 교란이 빈번한 시기가 끝난 직후인 11월 엽체의 생존률은 각각 48%와 42%로 큰열매모자반 이식군이 약간 높은 것으로 분석되었다(Fig. 3B). 이 결과는 원추상 부착기 고정 방식이 수지상 가근 고정방식과 최소한 동등하거나 우수한 해조류 이식방법이라는 것을 보여주었다. 이 연구에서 주목할 점은 큰열매모자반 단독 이식 그룹, 감태 단독 이식 그룹 및 두 분류군을 1:1로 혼합하여 이식한 그룹의 생존률 차이에 있다. 이식 초기 태풍에 의해 교란을 받는 기간(8~10월)을 포함하여 이식 1년 이상 지난 시점에서도 지속적으로 혼합 이식 그룹에서 단독 이식 그룹보다 높은 생존률을 나타내었다(Fig. 3B). 연구진이 수행하였던 사전 실험에서도 이와 유사한 결과가 나타났고(JGEC 2017), 향후 반복실험을 통해 결과를 다각도로 분석할 필요가 있다.
이식된 감태와 큰열매모자반 엽체의 생장은 이식시점인 8월부터 11월 동안 비교적 완만하게 진행되다가 수온이 15℃ 이하로 급격히 하강하는 1월부터 생장이 가속화되는 것으로 나타났으며, 이는 감태(Serisawa et al. 2002)와 큰열매모자반(Murase and Kito 1998; Ko et al. 2020)을 대상으로 실시한 이전 연구와 유사하였다. 또한, Serisawa et al. (2002)은 감태 부착기 발달이 이식 초기부터 5개월 동안 활발하게 이루어 진 후 정체기를 겪다가 이듬해 11월에서 2월 사이에 다시 활발한 생장기를 나타낸다고 하였는데, 이번 연구에서도 이식 초기인 가을부터 겨울 사이에 부착기가 활발하게 발달함을 확인하였다(Fig. 6A-C). 큰열매모자반의 부착기 또한 이식 후 1개월 동안 부착면의 상처가 아무는 현상이 관찰되었고, 그 후 겨울 동안 부착면이 기질을 따라 확장되는 양상을 보였다(Fig. 6D-F). 감태 및 큰열매모자반 두 분류군 모두 부착기는 엽체의 생장이 더딘 가을부터 초겨울 동안 왕성하게 발달함을 나타낸다. 따라서, 엽체의 생장과 부착기의 발달 추이를 고려하였을 때, 이 두 종의 대형 갈조류 이식 적기는 엽체의 크기가 작아 파도 등 물리적 교란에 대한 영향이 적고 부착기의 발달이 활발한 늦여름부터 초겨울이라는 것을 시사한다. 그러나 제주도의 경우 6월부터 10월까지는 태풍에 의한 교란이 빈번하고, 감태를 포함한 대형 갈조류를 섭식하는 것으로 알려진 아열대성 조식동물 독가시치(Siganus fuscescens)가 왕성하게 먹이활동을 하는 시기이므로, 이 시기에는 해조류 이식을 연기하거나 조식방지를 위한 조치가 필요하다(Nimura et al. 2007).
해조류 이식 후 약 2년이 경과한 2020년 6월, 이식된 해조류들의 생육 여부를 확인하였다. 감태와 큰열매모자반 개체군은 대부분 안정적으로 생육하고 있었고, 큰열매모자반 일부 개체군은 생식을 위한 소지(branchlet)를 무성하게 형성하였다. 인공기질의 상단면에는 새로 유입된 어린 감태(Fig. 7D)와 큰열매모자반(Fig. 7E)이 다수 확인되었다. 감태의 경우 실험장소 주변이 감태 우점군락이므로 어린 감태가 이식된 감태에 의해 재생산된 것인지는 검증이 필요하다. 그러나, 큰열매모자반은 주변 반경 50 m 이내에 자연 개체군이 관찰되지 않았으므로 새로운 유체는 본 실험의 결과 재생산된 개체군일 가능성이 높다.
이 실험은 자연 개체군을 채집하여 이식한 실험이기 때문에 이 방법을 지속적으로 사용할 경우 대량 채집으로 인한 자생 개체군 훼손이 우려된다. 따라서 인공적인 대량 생산 기법이 확립되기 전까지 한시적으로 사용하는 것이 바람직하다. 이 연구에서 큰열매모자반의 영양번식 가능성을 탐색하고자 줄기, 가지 및 정단부에 대해 동일한 방법으로 부착시험을 하였으나, 부착기가 성공적으로 발달하지 못하고 이식 1개월 후 모두 탈락하였다. 이를 보완하기 위한 방안으로 보다 다양한 방법의 영양번식 및 이식기술에 대한 연구가 필요하다(Yoshida et al. 1999). 이와 더불어, 인공기질 위에 이식한 해조류의 효과적인 재생산 요인을 탐색한 후, 재생산된 개체군을 다른 인공기질에 이식 적용하는 방법 즉, seed-bank를 조성하는 방안도 적극적으로 고려해야 한다.
결론적으로, 첫째 에폭시 수지와 순간접착제를 이용한 이식방법은 다년생 모자반류로 대표되는 원추상 부착기를 갖는 해조류의 효과적인 이식을 가능하게 하였다. 둘째, 해중림 조성을 위한 이식 대상종의 종다양성을 보다 높일 수 있는 방법을 제시하였다. 셋째, 엽상형 대형갈조 위주의 해중림 조성에서 다년생 모자반류를 혼합하여 이식함으로써 이식 해조류의 생존률이 상승하는 긍정적 효과를 도출하였다. 마지막으로 감태와 큰열매모자반의 이식 적기는 엽체 및 부착기 발달 양상, 태풍 영향 및 조식동물의 활동시기를 고려했을 때 가을부터 초겨울이 가장 적합한 시기인 것으로 판단된다.
